1 a Core on the Edge: an Urban Bird Assemblage on the Atlantic Margin
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
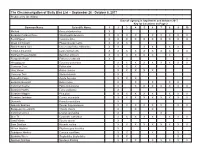
The Circumnavigation of Sicily Bird List -- September 26
The Circumnavigation of Sicily Bird List -- September 26 - October 8, 2017 Produced by Jim Wilson Date of sighting in September and October 2017 Key for Locations on Page 2 Common Name Scientific Name 1 2 3 4 5 6 7 8 9 10 Mallard Anas platyrhynchos X Eurasian Collared Dove Streptopelia decaocto X X X X X X X X X Feral Pigeon Columba livia X X X X X X X X X X Great Cormorant Phalacrocorax carbo X X X Black-headed Gull Chroicocephalus ridibundus X X X X X X Yellow-legged Gull Larus michahellis X X X X X X X X X X Northern House Martin Delichon urbicum X X X X X X X European Robin Erithacus rubecula X X Woodpigeon Columba palumbus X X X X X X X X X Common Coot Fulica atra X X X Grey Heron Ardea cinerea X X X X X Common Tern Sterna hirundo X Bonnelli's Eagle Aquila fasciata X X X Eurasian Buzzard Buteo buteo X X X X X Common Kestrel Falco tinnunculus X X X X X X X X Eurasian Hobby Falco subbuteo X Eurasian Magpie Pica pica X X X X X X X X Eurasian Jackdaw Corvus monedula X X X X X X X X X Dunnock Prunella modularis X Spanish Sparrow Passer hispaniolensis X X X X X X X X European Greenfinch Chloris chloris X X X Common Linnet Linaria cannabina X X Blue Tit Cyanistes caeruleus X X X X X X X X Sand Martin Riparia riparia X Barn Swallow Hirundo rustica X X X X X X X X X Willow Warbler Phylloscopus trochilus X Subalpine Warbler Curruca cantillans X X X X Eurasian Wren Troglodytes troglodytes X X X Spotless Starling Spotless Starling X X X X X X X X Common Name Scientific Name 1 2 3 4 5 6 7 8 9 10 Sardinian Warbler Curruca melanocephala X X X X -

Best of the Baltic - Bird List - July 2019 Note: *Species Are Listed in Order of First Seeing Them ** H = Heard Only
Best of the Baltic - Bird List - July 2019 Note: *Species are listed in order of first seeing them ** H = Heard Only July 6th 7th 8th 9th 10th 11th 12th 13th 14th 15th 16th 17th Mute Swan Cygnus olor X X X X X X X X Whopper Swan Cygnus cygnus X X X X Greylag Goose Anser anser X X X X X Barnacle Goose Branta leucopsis X X X Tufted Duck Aythya fuligula X X X X Common Eider Somateria mollissima X X X X X X X X Common Goldeneye Bucephala clangula X X X X X X Red-breasted Merganser Mergus serrator X X X X X Great Cormorant Phalacrocorax carbo X X X X X X X X X X Grey Heron Ardea cinerea X X X X X X X X X Western Marsh Harrier Circus aeruginosus X X X X White-tailed Eagle Haliaeetus albicilla X X X X Eurasian Coot Fulica atra X X X X X X X X Eurasian Oystercatcher Haematopus ostralegus X X X X X X X Black-headed Gull Chroicocephalus ridibundus X X X X X X X X X X X X European Herring Gull Larus argentatus X X X X X X X X X X X X Lesser Black-backed Gull Larus fuscus X X X X X X X X X X X X Great Black-backed Gull Larus marinus X X X X X X X X X X X X Common/Mew Gull Larus canus X X X X X X X X X X X X Common Tern Sterna hirundo X X X X X X X X X X X X Arctic Tern Sterna paradisaea X X X X X X X Feral Pigeon ( Rock) Columba livia X X X X X X X X X X X X Common Wood Pigeon Columba palumbus X X X X X X X X X X X Eurasian Collared Dove Streptopelia decaocto X X X Common Swift Apus apus X X X X X X X X X X X X Barn Swallow Hirundo rustica X X X X X X X X X X X Common House Martin Delichon urbicum X X X X X X X X White Wagtail Motacilla alba X X -

Niche Analysis and Conservation of Bird Species Using Urban Core Areas
sustainability Article Niche Analysis and Conservation of Bird Species Using Urban Core Areas Vasilios Liordos 1,* , Jukka Jokimäki 2 , Marja-Liisa Kaisanlahti-Jokimäki 2, Evangelos Valsamidis 1 and Vasileios J. Kontsiotis 1 1 Department of Forest and Natural Environment Sciences, International Hellenic University, 66100 Drama, Greece; [email protected] (E.V.); [email protected] (V.J.K.) 2 Arctic Centre, University of Lapland, 96101 Rovaniemi, Finland; jukka.jokimaki@ulapland.fi (J.J.); marja-liisa.kaisanlahti@ulapland.fi (M.-L.K.-J.) * Correspondence: [email protected] Abstract: Knowing the ecological requirements of bird species is essential for their successful con- servation. We studied the niche characteristics of birds in managed small-sized green spaces in the urban core areas of southern (Kavala, Greece) and northern Europe (Rovaniemi, Finland), during the breeding season, based on a set of 16 environmental variables and using Outlying Mean Index, a multivariate ordination technique. Overall, 26 bird species in Kavala and 15 in Rovaniemi were recorded in more than 5% of the green spaces and were used in detailed analyses. In both areas, bird species occupied different niches of varying marginality and breadth, indicating varying responses to urban environmental conditions. Birds showed high specialization in niche position, with 12 species in Kavala (46.2%) and six species in Rovaniemi (40.0%) having marginal niches. Niche breadth was narrower in Rovaniemi than in Kavala. Species in both communities were more strongly associated either with large green spaces located further away from the city center and having a high vegetation cover (urban adapters; e.g., Common Chaffinch (Fringilla coelebs), European Greenfinch (Chloris Citation: Liordos, V.; Jokimäki, J.; chloris Cyanistes caeruleus Kaisanlahti-Jokimäki, M.-L.; ), Eurasian Blue Tit ( )) or with green spaces located closer to the city center Valsamidis, E.; Kontsiotis, V.J. -

Phylogeny of the Eurasian Wren Nannus Troglodytes
PLOS ONE RESEARCH ARTICLE Phylogeny of the Eurasian Wren Nannus troglodytes (Aves: Passeriformes: Troglodytidae) reveals deep and complex diversification patterns of Ibero-Maghrebian and Cyrenaican populations 1 2 3 4 1 Frederik AlbrechtID *, Jens Hering , Elmar Fuchs , Juan Carlos Illera , Flora Ihlow , 5 5 6 7 a1111111111 Thomas J. ShannonID , J. Martin CollinsonID , Michael Wink , Jochen Martens , a1111111111 Martin PaÈckert1 a1111111111 a1111111111 1 Museum of Zoology, Senckenberg Natural History Collections Dresden, Senckenberg|Leibniz Institution for Biodiversity and Earth System Research, Dresden, Saxony, Germany, 2 Verein SaÈchsischer Ornithologen e. a1111111111 V., Limbach-Oberfrohna, Saxony, Germany, 3 Verein SaÈchsischer Ornithologen e.V., Weimar, Thuringia, Germany, 4 Research Unit of Biodiversity (UO-CSIC-PA), Oviedo University, Asturias, Spain, 5 School of Medicine, Medical Sciences and Nutrition, University of Aberdeen, Aberdeen, Scotland, United Kingdom, 6 Institute of Pharmacy and Molecular Biotechnology, Heidelberg University, Heidelberg, Baden- WuÈrttemberg, Germany, 7 Institute of Organismic and Molecular Evolution, Johannes Gutenberg University Mainz, Mainz, Rhineland-Palatinate, Germany OPEN ACCESS Citation: Albrecht F, Hering J, Fuchs E, Illera JC, * [email protected] Ihlow F, Shannon TJ, et al. (2020) Phylogeny of the Eurasian Wren Nannus troglodytes (Aves: Passeriformes: Troglodytidae) reveals deep and complex diversification patterns of Ibero- Abstract Maghrebian and Cyrenaican populations. PLoS The Mediterranean Basin represents a Global Biodiversity Hotspot where many organisms ONE 15(3): e0230151. https://doi.org/10.1371/ journal.pone.0230151 show high inter- and intraspecific differentiation. Extant phylogeographic patterns of terres- trial circum-Mediterranean faunas were mainly shaped through Pleistocene range shifts and Editor: èukasz Kajtoch, Institute of Systematics and Evolution of Animals Polish Academy of range fragmentations due to retreat into different glacial refugia. -

England Birds and Gardens 2013 Trip
Eagle-Eye England Birds and Gardens 2013 Trip May 7th – 18th 2013 Bird List Graylag Goose (Anser anser) Dark-bellied Brent Goose (Branta bernicla bernicla)] Canada Goose (Branta canadensis) Mute Swan (Cygnus olor) Egyptian Goose (Alopochen aegyptiaca) Common Shelduck (Tadorna tadorna) Mandarin Duck (Aix galericulata) Gadwall (Anas strepera) Eurasian Wigeon (Anas penelope) Mallard (Anas platyrhynchos) Northern Shoveler (Anas clypeata) Green-winged Teal (Eurasian) (Anas crecca crecca/nimia) Red-crested Pochard (Netta rufina) Common Pochard (Aythya ferina) Tufted Duck (Aythya fuligula) Common/Northern Eider (Somateria mollissima mollissima) Common Scoter (Melanitta nigra) Red-legged Partridge (Alectoris rufa) Gray Partridge (Perdix perdix) Ring-necked Pheasant (Phasianus colchicus) Little Grebe (Tachybaptus ruficollis) Great Crested Grebe (Podiceps cristatus) Northern Fulmar (Fulmarus glacialis) Northern Gannet (Morus bassanus) Great Cormorant (Phalacrocorax carbo) Gray Heron (Ardea cinerea) Little Egret (Egretta garzetta) Eurasian Spoonbill (Platalea leucorodia) Western Marsh-Harrier (Circus aeruginosus) Eurasian Sparrowhawk (Accipiter nisus) Common Buzzard (Buteo buteo) Water Rail (Rallus aquaticus) Eurasian Moorhen (Gallinula chloropus) Eurasian Coot (Fulica atra) Common Crane (Grus grus) Northern Lapwing (Vanellus vanellus) Grey/Black-bellied Plover (Pluvialis squatarola) Common Ringed Plover (Charadrius hiaticula) Little Ringed Plover (Charadrius dubius) Eurasian Oystercatcher (Haematopus ostralegus) Pied Avocet (Recurvirostra avosetta) -

Troglodytidae Species Tree
Troglodytidae I Rock Wren, Salpinctes obsoletus Canyon Wren, Catherpes mexicanus Sumichrast’s Wren, Hylorchilus sumichrasti Nava’s Wren, Hylorchilus navai Salpinctinae Nightingale Wren / Northern Nightingale-Wren, Microcerculus philomela Scaly-breasted Wren / Southern Nightingale-Wren, Microcerculus marginatus Flutist Wren, Microcerculus ustulatus Wing-banded Wren, Microcerculus bambla ?Gray-mantled Wren, Odontorchilus branickii Odontorchilinae Tooth-billed Wren, Odontorchilus cinereus Bewick’s Wren, Thryomanes bewickii Carolina Wren, Thryothorus ludovicianus Thrush-like Wren, Campylorhynchus turdinus Stripe-backed Wren, Campylorhynchus nuchalis Band-backed Wren, Campylorhynchus zonatus Gray-barred Wren, Campylorhynchus megalopterus White-headed Wren, Campylorhynchus albobrunneus Fasciated Wren, Campylorhynchus fasciatus Cactus Wren, Campylorhynchus brunneicapillus Yucatan Wren, Campylorhynchus yucatanicus Giant Wren, Campylorhynchus chiapensis Bicolored Wren, Campylorhynchus griseus Boucard’s Wren, Campylorhynchus jocosus Spotted Wren, Campylorhynchus gularis Rufous-backed Wren, Campylorhynchus capistratus Sclater’s Wren, Campylorhynchus humilis Rufous-naped Wren, Campylorhynchus rufinucha Pacific Wren, Nannus pacificus Winter Wren, Nannus hiemalis Eurasian Wren, Nannus troglodytes Zapata Wren, Ferminia cerverai Marsh Wren, Cistothorus palustris Sedge Wren, Cistothorus platensis ?Merida Wren, Cistothorus meridae ?Apolinar’s Wren, Cistothorus apolinari Timberline Wren, Thryorchilus browni Tepui Wren, Troglodytes rufulus Troglo dytinae Ochraceous -

Algerian Nuthatch Twitch (And a Bit of Tunesia) 05 Until 11-05-2018
ALGERIAN NUTHATCH TWITCH (AND A BIT OF TUNESIA) 05 UNTIL 11-05-2018 Max Berlijn, Epen, The Netherlands, [email protected] 1 Legenda: *AAAA = New (Holarctic) Species (2) AAAAA = Good species for the trip or for me (because not often seen) The list order is conformed the WP checklist with the recent changes on the splitting and lumping issue mainly based on publications in the “important magazines”. Subspecies is only mentioned when thought to be important and really visible in the field. The totals of birds per species are just a total of the birds I saw to give an idea how many of a species you encounter during a trip. Only species seen are mentioned, and heard when no lifer. On this trip the weather was good with no rain, mostly sunny with (dry) temperatures just above the 20 C and often no wind accept on the first day at Cap Bon. Cap Bon on a very windy day The breeding site of the first visited pair of Algerian Nuthatches Itinerary 05-05 Flew from Frankfurt to Tunis and met the group and the local guide Mohamed Ali Dakhli (+21621817717, [email protected] ) around 18.00 hours and drove to Cap Bon. 06-05 Birded around El Haouaria and Cap Bon all day. There was a very strong NW wind force 7. 07-05 Birded again around El Haouaria, with a three our pelagic from the cape. Later that day we drove to Tabarka near the Algerian border. Here we handed in our bins. 08-05 Drove from Tabarka to Jijel in Algeria. -

Conspecific Nest Aggression of The
CONSPECIFIC NEST AGGRESSION OF THE PACIFIC WREN ON VANCOUVER ISLAND, BRITISH COLUMBIA ANN NIGHTINGALE and RON MELCER, JR., Rocky Point Bird Observatory, Suite #170, 1581-H Hillside Avenue, Victoria, British Columbia, Canada V8T 2C1; [email protected] ABSTRACT: Five of the ten wren species in North America are known to destroy nests of conspecifics. These include the Cactus Wren (Campylorhynchus brunneica- pillus), Bewick’s Wren (Thryomanes bewickii), Sedge Wren (Cistothorus platensis), Marsh Wren (Cistothorus palustris), and House Wren (Troglodytes aedon). However, none of the Winter Wren complex, recently split as the Winter Wren (Troglodytes hiemalis), Pacific Wren (T. pacificus), and Eurasian Wren (T. troglodytes), have been documented to do so in experiments or by observation of natural behavior. Here we present a detailed chronology of a nesting of the Pacific Wren—the first report of conspecific nest aggression in the Winter Wren complex. On 15 May 2011, in Victoria, British Columbia, Canada, a Pacific Wren approached another’s nest under video surveillance and removed two 9-day-old chicks. The nonparental adult returned to the nest, apparently attempting to kill and/or and remove the remaining two chicks, several times over 4.75 hours but was not successful. Although our findings are limited to a single event, they are consistent with those of other wrens. Birds are known to destroy the nests and eggs or remove the young from nests of other species, as well as conspecifics, to reduce competition for nests, food, perches, and, in polygynous species, for the male’s parental care (Fox 1975, Verner 1975, Picman 1977a, Jones 1982). -

Wild Norway & Svalbard WILDLIFE SIGHTINGS (Birds & Mammals
Wild Norway & Svalbard WILDLIFE SIGHTINGS (Birds & Mammals) MAMMALS May June 19 20 21 22 23 24 25 26 27 28 29 30 31 1 Harbor Seal Phoca vitulina X X X European Otter Lutra lutra X X Arctic Hare Lepus arcticus X Reindeer Rangifer tarandus X X X X Harp Seal Pagophilus groenlandicus X Humpback Whale Megaptera novaeangliae X Ringed Seal Pusa hispida X X X X Walrus Odobenus rosmarus X X X Polar Bear Ursus maritimus X X Minke Whale Balaenoptera acutorostrata X Beluga (Whale) Delphinapterus ieucas X Blue Whale Balaenoptera musculus X BIRDS Meadow Pipit Anthus pratensis X X X X X X X X Skylark Alauda arvensis X X European Greenfinch Chloris chloris X X X X Common Redpoll Acanthis flammea X X X Common Linnet Linaria cannabina X Common Chaffinch Fringilla coelebs X X H H House Sparrow Passer domesticus X X X X X X X Common Starling Sturnus vulgaris X X X X X X X Hooded Crow Corvus cornix X X X X X X X Eurasian Magpie Pica pica X X X X X X Eurasian Blue Tit Cyanistes caeruleus X May June 19 20 21 22 23 24 25 26 27 28 29 30 31 1 Coal Tit Periparus ater X Great Tit Parus major H X Common Chiffchaff Phylloscopus collybita X X H H Willow Warbler Phylloscopus trochilus X X H X X X X Eurasian Blackcap Sylvia atricapilla H Northern Wheatear Oenanthe oenanthe X X X X X Common Blackbird Turdus merula X X Fieldfare Turdus pilaris X X X X Redwing Turdus iliacus X H X X X X X European Robin Charadrius hiaticula X Dunnock Prunella modularis X Eurasian Wren Troglodytes troglodytes X H H H Barn Swallow Hirundo rustica X X X X X X Sand Martin/Bank Riparia riparia -

The Historic England Zooarchaeology Reference Collection: Description and List
THE HISTORIC ENGLAND ZOOARCHAEOLOGY REFERENCE COLLECTION: DESCRIPTION AND LIST Polydora Baker, Eva Fairnell, Fay Worley SUMMARY This document lists the current holdings of the Historic England zooarchaeology reference collection held at Fort Cumberland, Portsmouth, UK. A total of 3308 specimens is listed, of which 2387 are complete skeletons. These include approximately 115 species of mammal, 220 species of bird, 9 species of reptile, 5 species of amphibian and 55 species of fish. The vertebrate skeleton reference collection is available for consultation: please refer to our access policy (http://www.historicengland.org.uk/research/approaches/research methods/Archaeology/zooarchaeology/). ACKNOWLEDGEMENTS The original collection list and description were compiled by Elaine Corke, Simon Davis and Sebastian Payne, and subsequently curated by Polydora Baker, Fay Worley, Mick Revill and Eva Fairnell. Complete specimen data recorded on individual paper records were added to the database by Gemma Ayton. We are grateful to Manny Lopez and Hugh Corley for IT support. COLLECTION LOCATION Historic England, Fort Cumberland, Fort Cumberland Rd, Portsmouth PO4 9LD, UK CONTACT DETAILS Zooarchaeology, Environmental Studies, Historic England, Fort Cumberland, Fort Cumberland Rd, Portsmouth PO4 9LD, UK Polydora Baker, Tel. 02392 856774. E-mail: [email protected] Fay Worley, Tel.: 02392 856789. E-mail: [email protected] 1 Historic England Zooarchaeology Reference Collection web list July 2020 INTRODUCTION The role of the Historic England Research Department vertebrate skeleton reference collection is to: • enable the identification and interpretation of animal remains from archaeological sites in England • provide comparative material for a long-term study of the development of the ‘traditional’ British domesticated animals • facilitate research on the biogeography of extinct and introduced species. -

Trip Report June 21 – July 2, 2017 | Written by Guide Gerard Gorman
Austria & Hungary: Trip Report June 21 – July 2, 2017 | Written by guide Gerard Gorman With Guide Gerard Gorman and Participants: Jane, Susan, and Cynthia and David This was the first Naturalist Journeys tour to combine Austria and Hungary, and it was a great success. We visited and birded some wonderfully diverse habitats and landscapes in 12 days — the forested foothills of the Alps in Lower Austria, the higher Alpine peak of Hochkar, the reed beds around Lake Neusiedler, the grasslands and wetlands of the Seewinkel in Austria and the Hanság in Hungary, the lowland steppes and farmlands of the Kiskunság east of the Danube, and finally the pleasant rolling limestone hills of the Bükk in north-east Hungary. In this way, we encountered diverse and distinct habitats and therefore birds, other wildlife, and flora, everywhere we went. We also stayed in a variety of different family-run guesthouses and lodges and sampled only local cuisine. As we went, we also found time to take in some of the rich history and culture of these two countries. Wed., June 21, 2017 We met up at Vienna International Airport at noon and were quickly on our way eastwards into Lower Austria. It was a pleasant sunny day and the traffic around the city was a little heavy, but we were soon away from the ring-road and in rural Austria. Roadside birds included both Carrion and Hooded Crows, several Eurasian Kestrels, and Barn Swallows zoomed overhead. Near Hainfeld we drove up a farm road, stopped for a picnic, and did our first real birding. -

A Checklist of Birds of Britain (9Th Edition)
Ibis (2018), 160, 190–240 doi: 10.1111/ibi.12536 The British List: A Checklist of Birds of Britain (9th edition) CHRISTOPHER J. MCINERNY,1,2 ANDREW J. MUSGROVE,1,3 ANDREW STODDART,1 ANDREW H. J. HARROP1 † STEVE P. DUDLEY1,* & THE BRITISH ORNITHOLOGISTS’ UNION RECORDS COMMITTEE (BOURC) 1British Ornithologists’ Union, PO Box 417, Peterborough PE7 3FX, UK 2School of Life Sciences, University of Glasgow, Glasgow G12 8QQ, UK 3British Trust for Ornithology, The Nunnery, Thetford, IP24 2PU, UK Recommended citation: British Ornithologists’ Union (BOU). 2018. The British List: a Checklist of Birds of Britain (9th edition). Ibis 160: 190–240. and the Irish Rare Birds Committee are no longer INTRODUCTION published within BOURC reports. This, the 9th edition of the Checklist of the Birds The British List is under continuous revision by of Britain, referred to throughout as the British BOURC. New species and subspecies are either List, has been prepared as a statement of the status added or removed, following assessment; these are of those species and subspecies known to have updated on the BOU website (https://www.bou. occurred in Britain and its coastal waters (Fig. 1). org.uk/british-list/recent-announcements/) at the time It incorporates all the changes to the British List of the change, but only come into effect on the List up to and including the 48th Report of the British on publication in a BOURC report in Ibis. A list of Ornithologists’ Union Records Committee the species and subspecies removed from the British (BOURC) (BOU 2018), and detailed in BOURC List since the 8th edition is shown in Appendix 1.