From Switzerland
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Theropod Composition of Early Late Cretaceous Faunas from Central
CORE Metadata, citation and similar papers at core.ac.uk Provided by Repository of the Academy's Library 1 Feeding related characters in basal pterosaurs: implications for jaw mechanism, dental function and diet RH: Feeding related characters in pterosaurs Attila Ősi A comparative study of various feeding related features in basal pterosaurs reveals a significant change in feeding strategies during the early evolutionary history of the group. These features are related to the skull architecture (e.g. quadrate morphology and orientation, jaw joint), dentition (e.g. crown morphology, wear patterns), reconstructed adductor musculature, and postcranium. The most basal pterosaurs (Preondactylus, dimorphodontids and anurognathids) were small bodied animals with a wing span no greater than 1.5 m, a relatively short, lightly constructed skull, straight mandibles with a large gape, sharply pointed teeth and well developed external adductors. The absence of extended tooth wear excludes complex oral food processing and indicates that jaw closure was simply orthal. Features of these basalmost forms indicate a predominantly insectivorous diet. Among stratigraphically older but more derived forms (Eudimorphodon, Carniadactylus, Caviramus) complex, multicusped teeth allowed the consumption of a wider variety of prey via a more effective form of food processing. This is supported by heavy dental wear in all forms with multicusped teeth. Typical piscivorous forms occurred no earlier than the Early Jurassic, and are characterized by widely spaced, enlarged procumbent teeth forming a fish grab and an anteriorly inclined quadrate that permitted only a relatively small gape. In addition, the skull became more elongate and body size 2 increased. Besides the dominance of piscivory, dental morphology and the scarcity of tooth wear reflect accidental dental occlusion that could have been caused by the capturing or seasonal consumption of harder food items. -

240 Delle - Boncourt - Porrentruy - Delémont État: 15
ANNÉE HORAIRE 2021 240 Delle - Boncourt - Porrentruy - Delémont État: 15. Octobre 2020 RE S3 RE S3 RE S3 RE S3 RE S3 RE 18159 17319 18161 17323 18163 17327 18165 17331 18167 17335 18169 [Meroux TGV] [Meroux TGV] Meroux TGV Meroux TGV dép. 06 03 08 03 09 03 Delle arr. 06 18 08 18 09 18 Delle 05 17 06 20 07 19 08 20 09 20 Boncourt 05 20 06 22 07 21 08 22 09 22 Buix 05 22 06 24 07 23 08 24 09 24 Grandgourt 05 24 07 25 Courtemaîche 05 28 06 30 07 30 08 30 09 30 Courchavon 05 30 06 32 07 32 08 32 09 32 Porrentruy 05 35 06 37 07 37 08 37 09 37 Bonfol dép. 05 52 06 22 06 52 07 22 07 43 08 22 09 22 Porrentruy arr. 06 07 06 37 07 07 07 37 07 58 08 37 09 37 Porrentruy 04 36 05 08 05 40 06 10 06 40 07 10 07 40 08 10 08 40 09 10 09 40 Courgenay 04 41 05 15 05 45 06 15 06 45 07 15 07 45 08 15 08 45 09 15 09 45 St-Ursanne 04 45 05 19 05 49 06 19 06 49 07 19 07 49 08 19 08 49 09 19 09 49 Glovelier 04 51 05 25 05 55 06 25 06 55 07 25 07 55 08 25 08 55 09 25 09 55 Saignelégier dép. 06 20 06 50 07 46 08 46 Glovelier arr. -

La Tuile, Un Objet Banal ? Levons Les Yeux
Parallèlement à sa production de vaisselle En Ajoie, la tuile apparaît à l’époque romaine. Un grand merci à nos sponsors et donateurs qui réfractaire, Bonfol a eu plusieurs tuileries. La plus Le nom latin tegula vient du verbe tegere qui ont permis de couvrir les frais de l’exposition. Sans ancienne apparaît vers 1820. La tuilerie Fattet, signifie couvrir et qui a donné « tuile » et « toit » en eux... quelle tuile ! créée vers 1883 à l’extérieur de la localité, a joué français. Après une éclipse, la tuile réapparaît au un rôle pionnier dans la mécanisation de la pro- Bas Moyen Âge sous la forme de la petite tuile duction jusqu’à sa disparition définitive en 1919. plate adaptée aux toits pentus. Vers la fin du 19e siècle, le procédé de production mécanique mis au La terre cuite architecturale point par les frères Gilardoni à Altkirch en 1841 est en Ajoie à travers les âges peu à peu mis en œuvre. Boulangerie L’Ami du Pain, Bonfol Louis Bélet SA, Vendlincourt Pius Borer, électricité, Bonfol A+C Corbat SA, industrie du bois, Vendlincourt Cosendey SA, construction, Bonfol Denner Bonmag Sàrl, Bonfol La tuile, un objet banal ? Filipetto Constructions, Vendlincourt Restaurant du Grütli, Bonfol Levons les yeux sur la variété Moderna SA, blanchisserie, Bonfol Technopoli SA, terminaison horlogère, Bonfol des couvertures protectrices Charpentes en bois Babey Sàrl, Boncourt de nos maisons. Charpente et couverture Maillard Philippe, Asuel Tuiles mécaniques de Bonfol. Un toit de Bonfol. Entreprise de construction Marcel Pheulpin, Bonfol Ouvert de mars à octobre La tuile est un élément architectural pesant et Outre l’argile, le tuilier n’a besoin que d’eau, Bienvenue au Musée de la poterie de Bonfol les 1er et 3e dimanches du mois résistant au temps et aux intempéries, mais de grands lieux de séchage et de stockage, de Autoroute A16 direction Porrentruy, sortie Courge- de 14 h 00 à 17 h 00 pas aux chocs. -

On Two Pterosaur Humeri from the Tendaguru Beds (Upper Jurassic, Tanzania)
“main” — 2009/10/20 — 22:40 — page 813 — #1 Anais da Academia Brasileira de Ciências (2009) 81(4): 813-818 (Annals of the Brazilian Academy of Sciences) ISSN 0001-3765 www.scielo.br/aabc On two pterosaur humeri from the Tendaguru beds (Upper Jurassic, Tanzania) FABIANA R. COSTA and ALEXANDER W.A. KELLNER Museu Nacional, Universidade Federal do Rio de Janeiro, Departamento de Geologia e Paleontologia Quinta da Boa Vista s/n, São Cristóvão, 20940-040 Rio de Janeiro, RJ, Brasil Manuscript received on August 17, 2009; accepted for publication on September 30, 2009; contributed by ALEXANDER W.A. KELLNER* ABSTRACT Jurassic African pterosaur remains are exceptionally rare and only known from the Tendaguru deposits, Upper Jurassic, Tanzania. Here we describe two right humeri of Tendaguru pterosaurs from the Humboldt University of Berlin: specimens MB.R. 2828 (cast MN 6661-V) and MB.R. 2833 (cast MN 6666-V). MB.R. 2828 consists of a three- dimensionally preserved proximal portion. The combination of the morphological features of the deltopectoral crest not observed in other pterosaurs suggests that this specimen belongs to a new dsungaripteroid taxon. MB.R. 2833 is nearly complete, and because of a long and round proximally placed deltopectoral crest it could be referred to the Archaeopterodactyloidea. It is the smallest pterosaur from Africa and one of the smallest flying reptiles ever recorded. These specimens confirm the importance of the Tendaguru deposits for the Jurassic pterosaur record. This potential, however, has to be fully explored with more field work. Key words: Tendaguru, Tanzania, Africa, Upper Jurassic, Pterosauria. INTRODUCTION in providing isolated remains up to now (Kellner and Mader 1997, Wellnhofer and Buffetaut 1999, Mader Africa shows a great potential for pterosaur material and Kellner 1999). -

Analyzing Pterosaur Ontogeny and Sexual Dimorphism with Multivariate Allometry Erick Charles Anderson [email protected]
Marshall University Marshall Digital Scholar Theses, Dissertations and Capstones 2016 Analyzing Pterosaur Ontogeny and Sexual Dimorphism with Multivariate Allometry Erick Charles Anderson [email protected] Follow this and additional works at: http://mds.marshall.edu/etd Part of the Animal Sciences Commons, Ecology and Evolutionary Biology Commons, and the Paleontology Commons Recommended Citation Anderson, Erick Charles, "Analyzing Pterosaur Ontogeny and Sexual Dimorphism with Multivariate Allometry" (2016). Theses, Dissertations and Capstones. 1031. http://mds.marshall.edu/etd/1031 This Thesis is brought to you for free and open access by Marshall Digital Scholar. It has been accepted for inclusion in Theses, Dissertations and Capstones by an authorized administrator of Marshall Digital Scholar. For more information, please contact [email protected], [email protected]. ANALYZING PTEROSAUR ONTOGENY AND SEXUAL DIMORPHISM WITH MULTIVARIATE ALLOMETRY A thesis submitted to the Graduate College of Marshall University In partial fulfillment of the requirements for the degree of Master of Science in Biological Sciences by Erick Charles Anderson Approved by Dr. Frank R. O’Keefe, Committee Chairperson Dr. Suzanne Strait Dr. Andy Grass Marshall University May 2016 i ii ii Erick Charles Anderson ALL RIGHTS RESERVED iii Acknowledgments I would like to thank Dr. F. Robin O’Keefe for his guidance and advice during my three years at Marshall University. His past research and experience with reptile evolution made this research possible. I would also like to thank Dr. Andy Grass for his advice during the course of the research. I would like to thank my fellow graduate students Donald Morgan and Tiffany Aeling for their support, encouragement, and advice in the lab and bar during our two years working together. -

Portrait De Territoire Nord Franche
Nord Franche-Comté - Canton du Jura Un territoire contrasté entre une partie française urbaine à forte densité et un côté suisse moins peuplé vec 330 000 habitants, le territoire « Nord Franche-Comté - Canton du Jura » est le plus peuplé des quatre territoires de coopération. Il comprend les agglomérations françaises de Belfort et de Montbéliard. En Suisse, deux pôles d’emploi importants, ceux de Delémont et de Porrentruy, attirent des actifs résidant dans les petites communes françaises situées le long de la frontière. L’industrie est bien implantée dans ce territoire, structurée autour de grands établissements relevant en grande partie de la fabrication de matériel de transport côté Afrançais, davantage diversifiée côté suisse. Démographie : une évolution contrastée Ce territoire se caractérise par de part et d’autre de la frontière des collaborations régulières et multiscalaires qui associent principalement le Canton du Jura, Le territoire « Nord Franche-Comté - Canton (14 100) et une vingtaine de communes de le Département du Territoire de Belfort, du Jura » est un territoire de 330 000 habitants plus de 2000 habitants. la Communauté de l’Agglomération situé dans la partie la moins montagneuse de Le contraste est fort avec la partie suisse, si- Belfortaine (CAB) et la Préfecture du l’Arc jurassien. De fait, il est densément peu- tuée dans le canton du Jura, qui compte 105 Territoire de Belfort. plé, avec 226 habitants au km² alors que la habitants au km² et comprend moins d’une di- De nombreuses réalisations sont à relever, moyenne de l’Arc jurassien se situe à 133 ha- zaine de communes dépassant les 2000 habi- notamment dans le domaine culturel bitants au km². -

Pterosaur Distribution in Time and Space: an Atlas 61
Zitteliana An International Journal of Palaeontology and Geobiology Series B/Reihe B Abhandlungen der Bayerischen Staatssammlung für Pa lä on to lo gie und Geologie B28 DAVID W. E. HONE & ERIC BUFFETAUT (Eds) Flugsaurier: pterosaur papers in honour of Peter Wellnhofer CONTENTS/INHALT Dedication 3 PETER WELLNHOFER A short history of pterosaur research 7 KEVIN PADIAN Were pterosaur ancestors bipedal or quadrupedal?: Morphometric, functional, and phylogenetic considerations 21 DAVID W. E. HONE & MICHAEL J. BENTON Contrasting supertree and total-evidence methods: the origin of the pterosaurs 35 PAUL M. BARRETT, RICHARD J. BUTLER, NICHOLAS P. EDWARDS & ANDREW R. MILNER Pterosaur distribution in time and space: an atlas 61 LORNA STEEL The palaeohistology of pterosaur bone: an overview 109 S. CHRISTOPHER BENNETT Morphological evolution of the wing of pterosaurs: myology and function 127 MARK P. WITTON A new approach to determining pterosaur body mass and its implications for pterosaur fl ight 143 MICHAEL B. HABIB Comparative evidence for quadrupedal launch in pterosaurs 159 ROSS A. ELGIN, CARLOS A. GRAU, COLIN PALMER, DAVID W. E. HONE, DOUGLAS GREENWELL & MICHAEL J. BENTON Aerodynamic characters of the cranial crest in Pteranodon 167 DAVID M. MARTILL & MARK P. WITTON Catastrophic failure in a pterosaur skull from the Cretaceous Santana Formation of Brazil 175 MARTIN LOCKLEY, JERALD D. HARRIS & LAURA MITCHELL A global overview of pterosaur ichnology: tracksite distribution in space and time 185 DAVID M. UNWIN & D. CHARLES DEEMING Pterosaur eggshell structure and its implications for pterosaur reproductive biology 199 DAVID M. MARTILL, MARK P. WITTON & ANDREW GALE Possible azhdarchoid pterosaur remains from the Coniacian (Late Cretaceous) of England 209 TAISSA RODRIGUES & ALEXANDER W. -

Patinoire De Porrentruy Annonce Des Postes Vacants EFEJ À Bassecourt
N°43 I Janvier 2020 SERVICE DE L’ÉCONOMIE ET DE L’EMPLOI OFFICE RÉGIONAL DE PLACEMENT Patinoire de Porrentruy L’affaire des entreprises jurassiennes Annonce des postes vacants Premier bilan encourageant EFEJ à Bassecourt Une imprimante 3D pour la formation Photo : Stähelin partenar Dolci architectes Obligation d’annoncer les postes vacants – dès le 1er juillet 2018 Obligation d’annoncer les postes vacants – dès le 1er juillet 2018 Informations sur: Informations sur: travail.swiss Actualité travail.swiss Premier bilan positif pour l’OAPV L’obligation d’annoncer les postes vacants ( OAPV ), consécutive à l’initiative « Contre l’immigration de masse » acceptée le 9 février 2014, a été introduite voici 18 mois. Bilan et perspectives. Depuis mi-2018, cette mesure, dite L’industrie « préférence indigène light », oblige en première ligne les employeurs à annoncer leurs postes vacants en priorité aux Offices Durant la première année, régionaux de placement ( ORP ). Elle l’ORP-Jura a reçu 2468 s’applique aux genres de professions annonces d’emplois au to- les plus touchés par le chômage. Les tal. Mille quatre cent onze demandeurs d’emploi reçoivent ain- ( 57 % ) étaient obligatoires, si l’information prioritairement et en dont plus de 60 % concer- exclusivité durant 5 jours. Ils peuvent naient des postes fixes ou postuler avec une longueur d’avance, temporaires dans l’indus- alors que les ORP transmettent pa- trie, souvent annoncés par rallèlement des dossiers aux em- des agences privées de ployeurs. placement. Pas étonnant vu la structure de notre Les entreprises jouent le jeu économie et la conjoncture favorable, qui a prévalu Le Secrétariat d’État à l’économie durant la période examinée, et qui a siers suite à une prise d’emploi. -

New Perspectives on Pterosaur Palaeobiology
Downloaded from http://sp.lyellcollection.org/ by guest on September 24, 2021 New perspectives on pterosaur palaeobiology DAVID W. E. HONE1*, MARK P. WITTON2 & DAVID M. MARTILL2 1Queen Mary University of London, Mile End Road, London E14NS, UK 2School of Earth and Environmental Sciences, University of Portsmouth, Burnaby Road, Portsmouth PO1 3QL, UK *Correspondence: [email protected] Abstract: Pterosaurs were the first vertebrates to evolve powered flight and occupied the skies of the Mesozoic for 160 million years. They occurred on every continent, evolved their incredible pro- portions and anatomy into well over 100 species, and included the largest flying animals of all time among their ranks. Pterosaurs are undergoing a long-running scientific renaissance that has seen elevated interest from a new generation of palaeontologists, contributions from scientists working all over the world and major advances in our understanding of their palaeobiology. They have espe- cially benefited from the application of new investigative techniques applied to historical speci- mens and the discovery of new material, including detailed insights into their fragile skeletons and their soft tissue anatomy. Many aspects of pterosaur science remain controversial, mainly due to the investigative challenges presented by their fragmentary, fragile fossils and notoriously patchy fossil record. With perseverance, these controversies are being resolved and our understand- ing of flying reptiles is increasing. This volume brings together a diverse set of papers on numerous aspects of the biology of these fascinating reptiles, including discussions of pterosaur ecology, flight, ontogeny, bony and soft tissue anatomy, distribution and evolution, as well as revisions of their taxonomy and relationships. -
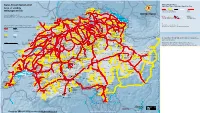
Swiss Travel System Map 2021
ai160326587010_STS-GB-Pass-S-21.pdf 1 21.10.20 09:37 Kruth Strasbourg | Paris Karlsruhe | Frankfurt | Dortmund | Hamburg | Berlin Stuttgart Ulm | München München Swiss Travel System 2021 Stockach Swiss Travel Pass Blumberg-Zollhaus Engen Swiss Travel Pass Youth | Swiss Travel Pass Flex Bargen Opfertshofen Überlingen Area of validity Seebrugg Beggingen Singen Ravensburg DEUTSCHLAND Radolfzell Schleitheim Hemmental Lines for unlimited travel (tunnel) Mulhouse Thayngen Mainau Geltungsbereich Meersburg Schaffhausen Ramsen Linien für unbegrenzte Fahrten (Tunnel) Zell (Wiesental) Wangen (Allgäu) Erzingen Oster- Neuhausen Stein a.R. Konstanz fingen Version/Stand/Etat/Stato:12.2020 (Baden) Rheinau Kreuzlingen Friedrichshafen Waldshut Due to lack of space not all lines are indicated. Subject to change. Marthalen Basel Weil a.R. Aus Platzgründen sind nicht alle Linien angegeben. Änderungen vorbehalten. Bad Zurzach Weinfelden Lines with reductions (50%, 1 25%) No reductions EuroAirport Riehen Koblenz Eglisau Frauenfeld Romanshorn Lindau Basel St.Johann Basel Möhlin Laufenburg Immenstadt Linien mit Vergünstigungen (50%, 1 25%) Keine Ermässigung Bad Bf Nieder- Stein-Säckingen Bülach Sulgen Arbon Basel Rheinfelden weningen Braunau Sonthofen Delle Pratteln Turgi Rorschach Bregenz Boncourt Ettingen Frick Brugg Zürich Bischofszell Rheineck Bonfol Liestal Baden Flughafen Winterthur Wil Rodersdorf Dornach Oberglatt Heiden St.Margrethen Aesch Gelterkinden Kloten Turbenthal St.Gallen Walzenhausen Roggenburg Wettingen Also valid for local public transport -

Cantons De L'arrondissement D'arras
- 2 - CANTON D’ARRAS NORD 29 emplacements ARRAS 8 emplacement(s) - Rue Jacques le Caron, Hôtel de Ville - Rue de Justice, école Anatole France - Allée des Rosati, école Georges Brassens - Rue Guinegatte, école Victor Hugo - Grande rue du Rivage, foyer Soleil - Office Culturel, Grand'Place - Rue du Dr Brassart, mur de clôture (limite de la place Foch) - Résidence Saint Michel (au milieu de la rue Saint Michel, sur les grilles devant la pharmacie) ATHIES 6 emplacement(s) - Place du Maréchal Foch - Rue Pierre Létienne - Grand'Rue - Rue Arthur Delobelle - Rue d'Arras - Mur du cimetière SAINT LAURENT BLANGY 5 emplacement(s) - Rue Laurent Gers n° 8, Hôtel de Ville - Rue de Versailles n° 50, école Lenglet - Rue de la Sambre n° 40, école Langevin - Rue d'Amboise n° 21, école Les Capucines - Rue du Général de Gaulle n° 26, centre culturel Vivot SAINT NICOLAS 10 emplacement(s) - Centre Bonne Humeur - Groupe scolaire Grenier - Rue Anatole France, face aux ateliers communaux - Rue Raoul Briquet, mur côté Poste - Rue du Zodiaque, le long de l'école Grenier - Rue de la Forge au Fer, clôture du stade de la Scarpe - Avenue Raoul Thibaut, face à la résidence de la Chasse - Rue Aristide Briand, le long du mur du centre Camille Corot - Place des Ecrins, bâtiment Le Pelvoux - Rue du Zodiaque face au hameau du Scorpion .../... - 3 - CANTON D’ARRAS OUEST 19 emplacements ARRAS 19 emplacement(s) - Place Quincaille, école Paul Bert - 16, Rue A. Briand , maison des Sociétés, - Rue Albert Camus, école Charles Péguy - Rue Charles Péguy n° 1, maison de service de proximité M.T. -

Redalyc.On Two Pterosaur Humeri from the Tendaguru Beds (Upper Jurassic, Tanzania)
Anais da Academia Brasileira de Ciências ISSN: 0001-3765 [email protected] Academia Brasileira de Ciências Brasil COSTA, FABIANA R.; KELLNER, ALEXANDER W.A. On two pterosaur humeri from the Tendaguru beds (Upper Jurassic, Tanzania) Anais da Academia Brasileira de Ciências, vol. 81, núm. 4, diciembre, 2009, pp. 813-818 Academia Brasileira de Ciências Rio de Janeiro, Brasil Available in: http://www.redalyc.org/articulo.oa?id=32713481017 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative “main” — 2009/10/20 — 22:40 — page 813 — #1 Anais da Academia Brasileira de Ciências (2009) 81(4): 813-818 (Annals of the Brazilian Academy of Sciences) ISSN 0001-3765 www.scielo.br/aabc On two pterosaur humeri from the Tendaguru beds (Upper Jurassic, Tanzania) FABIANA R. COSTA and ALEXANDER W.A. KELLNER Museu Nacional, Universidade Federal do Rio de Janeiro, Departamento de Geologia e Paleontologia Quinta da Boa Vista s/n, São Cristóvão, 20940-040 Rio de Janeiro, RJ, Brasil Manuscript received on August 17, 2009; accepted for publication on September 30, 2009; contributed by ALEXANDER W.A. KELLNER* ABSTRACT Jurassic African pterosaur remains are exceptionally rare and only known from the Tendaguru deposits, Upper Jurassic, Tanzania. Here we describe two right humeri of Tendaguru pterosaurs from the Humboldt University of Berlin: specimens MB.R. 2828 (cast MN 6661-V) and MB.R. 2833 (cast MN 6666-V). MB.R.