Assessment of the Pacific Cod Stock in the Eastern Bering Sea
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Fishery Management Plan for Groundfish of the Bering Sea and Aleutian Islands Management Area APPENDICES
FMP for Groundfish of the BSAI Management Area Fishery Management Plan for Groundfish of the Bering Sea and Aleutian Islands Management Area APPENDICES Appendix A History of the Fishery Management Plan ...................................................................... A-1 A.1 Amendments to the FMP ......................................................................................................... A-1 Appendix B Geographical Coordinates of Areas Described in the Fishery Management Plan ..... B-1 B.1 Management Area, Subareas, and Districts ............................................................................. B-1 B.2 Closed Areas ............................................................................................................................ B-2 B.3 PSC Limitation Zones ........................................................................................................... B-18 Appendix C Summary of the American Fisheries Act and Subtitle II ............................................. C-1 C.1 Summary of the American Fisheries Act (AFA) Management Measures ............................... C-1 C.2 Summary of Amendments to AFA in the Coast Guard Authorization Act of 2010 ................ C-2 C.3 American Fisheries Act: Subtitle II Bering Sea Pollock Fishery ............................................ C-4 Appendix D Life History Features and Habitat Requirements of Fishery Management Plan SpeciesD-1 D.1 Walleye pollock (Theragra calcogramma) ............................................................................ -

2020-2021 Statewide Commercial Groundfish Fishing Regulations
Alaska Department of Fish and Game 2020–2021 Statewide Commercial Groundfish Fishing Regulations This booklet contains regulations regarding COMMERCIAL GROUNDFISH FISHERIES in the State of Alaska. This booklet covers the period May 2020 through March 2021 or until a new book is available following the Board of Fisheries meetings. Note to Readers: These statutes and administrative regulations were excerpted from the Alaska Statutes (AS), and the Alaska Administrative Code (AAC) based on the official regulations on file with the Lieutenant Governor. There may be errors or omissions that have not been identified and changes that occurred after this printing. This booklet is intended as an informational guide only. To be certain of the current laws, refer to the official statutes and the AAC. Changes to Regulations in this booklet: The regulations appearing in this booklet may be changed by subsequent board action, emergency regulation, or emergency order at any time. Supplementary changes to the regulations in this booklet will be available on the department′s website and at offices of the Department of Fish and Game. For information or questions regarding regulations, requirements to participate in commercial fishing activities, allowable activities, other regulatory clarifications, or questions on this publication please contact the Regulations Program Coordinator at (907) 465-6124 or email [email protected] The Alaska Department of Fish and Game (ADF&G) administers all programs and activities free from discrimination based on race, color, national origin, age, sex, religion, marital status, pregnancy, parenthood, or disability. The department administers all programs and activities in compliance with Title VI of the Civil Rights Act of 1964, Section 504 of the Rehabilitation Act of 1973, Title II of the Americans with Disabilities Act of 1990, the Age Discrimination Act of 1975, and Title IX of the Education Amendments of 1972. -

Humboldt Bay Fishes
Humboldt Bay Fishes ><((((º>`·._ .·´¯`·. _ .·´¯`·. ><((((º> ·´¯`·._.·´¯`·.. ><((((º>`·._ .·´¯`·. _ .·´¯`·. ><((((º> Acknowledgements The Humboldt Bay Harbor District would like to offer our sincere thanks and appreciation to the authors and photographers who have allowed us to use their work in this report. Photography and Illustrations We would like to thank the photographers and illustrators who have so graciously donated the use of their images for this publication. Andrey Dolgor Dan Gotshall Polar Research Institute of Marine Sea Challengers, Inc. Fisheries And Oceanography [email protected] [email protected] Michael Lanboeuf Milton Love [email protected] Marine Science Institute [email protected] Stephen Metherell Jacques Moreau [email protected] [email protected] Bernd Ueberschaer Clinton Bauder [email protected] [email protected] Fish descriptions contained in this report are from: Froese, R. and Pauly, D. Editors. 2003 FishBase. Worldwide Web electronic publication. http://www.fishbase.org/ 13 August 2003 Photographer Fish Photographer Bauder, Clinton wolf-eel Gotshall, Daniel W scalyhead sculpin Bauder, Clinton blackeye goby Gotshall, Daniel W speckled sanddab Bauder, Clinton spotted cusk-eel Gotshall, Daniel W. bocaccio Bauder, Clinton tube-snout Gotshall, Daniel W. brown rockfish Gotshall, Daniel W. yellowtail rockfish Flescher, Don american shad Gotshall, Daniel W. dover sole Flescher, Don stripped bass Gotshall, Daniel W. pacific sanddab Gotshall, Daniel W. kelp greenling Garcia-Franco, Mauricio louvar -

Guide to the Coastal Marine Fishes of California
STATE OF CALIFORNIA THE RESOURCES AGENCY DEPARTMENT OF FISH AND GAME FISH BULLETIN 157 GUIDE TO THE COASTAL MARINE FISHES OF CALIFORNIA by DANIEL J. MILLER and ROBERT N. LEA Marine Resources Region 1972 ABSTRACT This is a comprehensive identification guide encompassing all shallow marine fishes within California waters. Geographic range limits, maximum size, depth range, a brief color description, and some meristic counts including, if available: fin ray counts, lateral line pores, lateral line scales, gill rakers, and vertebrae are given. Body proportions and shapes are used in the keys and a state- ment concerning the rarity or commonness in California is given for each species. In all, 554 species are described. Three of these have not been re- corded or confirmed as occurring in California waters but are included since they are apt to appear. The remainder have been recorded as occurring in an area between the Mexican and Oregon borders and offshore to at least 50 miles. Five of California species as yet have not been named or described, and ichthyologists studying these new forms have given information on identification to enable inclusion here. A dichotomous key to 144 families includes an outline figure of a repre- sentative for all but two families. Keys are presented for all larger families, and diagnostic features are pointed out on most of the figures. Illustrations are presented for all but eight species. Of the 554 species, 439 are found primarily in depths less than 400 ft., 48 are meso- or bathypelagic species, and 67 are deepwater bottom dwelling forms rarely taken in less than 400 ft. -
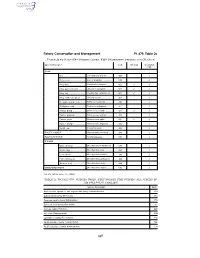
Species Codes: Fmp Prohibited Species and Cr Crab
Fishery Conservation and Management Pt. 679, Table 2c TABLE 2b TO PART 679—SPECIES CODES: FMP PROHIBITED SPECIES AND CR CRAB Species Description Code CR Crab Groundfish PSC CRAB Box ....................................... Lopholithodes mandtii .......... 900 ✓ Dungeness ........................... Cancer magister .................. 910 ✓ King, blue ............................. Paralithodes platypus .......... 922 ✓ ✓ King, golden (brown) ........... Lithodes aequispinus ........... 923 ✓ ✓ King, red .............................. Paralithodes camtshaticus ... 921 ✓ ✓ King, scarlet (deepsea) ....... Lithodes couesi .................... 924 ✓ Korean horsehair crab ......... Erimacrus isenbeckii ............ 940 ✓ Multispinus crab ................... Paralomis multispinus .......... 951 ✓ Tanner, Bairdi ...................... Chionoecetes bairdi ............. 931 ✓ ✓ Tanner, grooved .................. Chionoecetes tanneri ........... 933 ✓ Tanner, snow ....................... Chionoecetes opilio ............. 932 ✓ ✓ Tanner, triangle ................... Chionoecetes angulatus ...... 934 ✓ Verrilli crab ........................... Paralomis verrilli .................. 953 ✓ PACIFIC HALIBUT Hippoglossus stenolepis ...... 200 ✓ PACIFIC HERRING Family Clupeidae ................. 235 ✓ SALMON Chinook (king) ..................... Oncorhynchus tshawytscha 410 ✓ Chum (dog) .......................... Oncorhynchus keta .............. 450 ✓ Coho (silver) ........................ Oncorhynchus kisutch ......... 430 ✓ Pink (humpback) ................. -

1 CWU Comparative Osteology Collection, List of Specimens
CWU Comparative Osteology Collection, List of Specimens List updated November 2019 0-CWU-Collection-List.docx Specimens collected primarily from North American mid-continent and coastal Alaska for zooarchaeological research and teaching purposes. Curated at the Zooarchaeology Laboratory, Department of Anthropology, Central Washington University, under the direction of Dr. Pat Lubinski, [email protected]. Facility is located in Dean Hall Room 222 at CWU’s campus in Ellensburg, Washington. Numbers on right margin provide a count of complete or near-complete specimens in the collection. Specimens on loan from other institutions are not listed. There may also be a listing of mount (commercially mounted articulated skeletons), part (partial skeletons), skull (skulls), or * (in freezer but not yet processed). Vertebrate specimens in taxonomic order, then invertebrates. Taxonomy follows the Integrated Taxonomic Information System online (www.itis.gov) as of June 2016 unless otherwise noted. VERTEBRATES: Phylum Chordata, Class Petromyzontida (lampreys) Order Petromyzontiformes Family Petromyzontidae: Pacific lamprey ............................................................. Entosphenus tridentatus.................................... 1 Phylum Chordata, Class Chondrichthyes (cartilaginous fishes) unidentified shark teeth ........................................................ ........................................................................... 3 Order Squaliformes Family Squalidae Spiny dogfish ........................................................ -

Fishes-Of-The-Salish-Sea-Pp18.Pdf
NOAA Professional Paper NMFS 18 Fishes of the Salish Sea: a compilation and distributional analysis Theodore W. Pietsch James W. Orr September 2015 U.S. Department of Commerce NOAA Professional Penny Pritzker Secretary of Commerce Papers NMFS National Oceanic and Atmospheric Administration Kathryn D. Sullivan Scientifi c Editor Administrator Richard Langton National Marine Fisheries Service National Marine Northeast Fisheries Science Center Fisheries Service Maine Field Station Eileen Sobeck 17 Godfrey Drive, Suite 1 Assistant Administrator Orono, Maine 04473 for Fisheries Associate Editor Kathryn Dennis National Marine Fisheries Service Offi ce of Science and Technology Fisheries Research and Monitoring Division 1845 Wasp Blvd., Bldg. 178 Honolulu, Hawaii 96818 Managing Editor Shelley Arenas National Marine Fisheries Service Scientifi c Publications Offi ce 7600 Sand Point Way NE Seattle, Washington 98115 Editorial Committee Ann C. Matarese National Marine Fisheries Service James W. Orr National Marine Fisheries Service - The NOAA Professional Paper NMFS (ISSN 1931-4590) series is published by the Scientifi c Publications Offi ce, National Marine Fisheries Service, The NOAA Professional Paper NMFS series carries peer-reviewed, lengthy original NOAA, 7600 Sand Point Way NE, research reports, taxonomic keys, species synopses, fl ora and fauna studies, and data- Seattle, WA 98115. intensive reports on investigations in fi shery science, engineering, and economics. The Secretary of Commerce has Copies of the NOAA Professional Paper NMFS series are available free in limited determined that the publication of numbers to government agencies, both federal and state. They are also available in this series is necessary in the transac- exchange for other scientifi c and technical publications in the marine sciences. -

2013-2014 Commercial Finfish and Shellfish Fishing General
Alaska Department of Fish and Game 2013–2014 Commercial Finfish and Shellfish Fishing General Regulation & Statute Provisions This booklet contains statutes and general provisions regarding commercial fisheries in the State of Alaska. This booklet covers the period June 2013 through June 2014 or until the 2014 book is available, whichever occurs first. Note to Readers: These statutes and administrative regulations were excerpted from the Alaska Statutes and the Alaska Administrative Code (AAC) based on the official regulations on file with the Lieutenant Governor. There may be errors or omissions that have not been identified and changes that occurred after this printing. This booklet is intended as an informational guide only. To be certain of the current laws, refer to the official statutes and the AAC. Changes to Regulations in this booklet: The regulations appearing in this booklet may be changed by emergency regulation or emergency order at any time. Supplementary changes to the regulations in this booklet will be available at offices of the Department of Fish and Game. This publication was released by the Department of Fish and Game. The Alaska Department of Fish and Game (ADF&G) administers all programs and activities free from discrimination based on race, color, national origin, age, sex, religion, marital status, pregnancy, parenthood, or disability. The department administers all programs and activities in compliance with Title VI of the Civil Rights Act of 1964, Section 504 of the Rehabilitation Act of 1973, Title II of the Americans with Disabilities Act of 1990, the Age Discrimination Act of 1975, and Title IX of the Education Amendments of 1972. -

2019 Commercial Synopsis.Pdf
ATTENTION This publication describes the non-Indian commercial fishing regulations in condensed form for the convenience of the commercial harvester and is not intended to replace the laws or administrative rules. Oregon Revised Statutes (ORS) and Oregon Administrative Rules (OAR) are cited in parentheses to help the reader refer to the original sections that were condensed. For detailed information refer to these laws and rules. Unless otherwise specified, information covers laws and rules in effect at the start of the year and is subject to change during the year. Copies of specific laws and rules may be obtained from Oregon Department of Fish and Wildlife (ODFW) offices. The ORS and OAR are also available online (see pages 6 and 7) and in most public libraries. Commercial fishery laws not listed here are still applicable. Any person participating in any commercial fishery in Oregon must abide by all applicable state and federal laws and rules. Related Publications: 1. Federal Regulations Applying in the Exclusive Economic Zone (EEZ) from 3- 200 miles for the U.S. Commercial and Recreational Groundfish Fisheries off the Coasts of Washington, Oregon, and California. 2. Federal Regulations for West Coast Salmon Fisheries Applying in the Exclusive Economic Zone (3-200 miles) off the Coasts of Washington, Oregon, and California. 3. Current information relating to the EEZ for groundfish and salmon federal regulations are available at the website for the National Marine Fisheries Service (NMFS) West Coast Regional Office (WCR): www.westcoast.fisheries.noaa.gov -

Spatial and Temporal Variation in Tufted Puffin Fratercula Cirrhata Nestling Diet Quality and Growth Rates
Williams & Buck: Puffin diets and growth 41 SPATIAL AND TEMPORAL VARIATION IN TUFTED PUFFIN FRATERCULA CIRRHATA NESTLING DIET QUALITY AND GROWTH RATES CORY T. WILLIAMS & C. LOREN BUCK Department of Biological Sciences, University of Alaska Anchorage, Anchorage, Alaska, 99508, USA ([email protected]) Received 28 July 2009, accepted 15 February 2010 SUMMARY WILLIAMS, C.T. & BUCK, C.L. 2010. Spatial and temporal variation in Tufted Puffin Fratercula cirrhata nestling diet quality and growth rates. Marine Ornithology 38: 41–48. Seabirds have long been promoted as bio-indicators, because parameters such as reproductive success, nestling growth rates, and diet composition respond markedly to changes in food supply. Although such responses are often associated with broad-scale oceanographic phenomena, they are also influenced by processes that occur on much smaller spatial scales. We quantified Tufted Puffin Fratercula cirrhata nestling growth rates, apparent fledging success, and diets at several colonies in Chiniak Bay, Kodiak Island, Alaska from 2003-2005. We also measured the lipid content of forage fish fed to nestlings, because diet selection is expected to be influenced by prey quality. Although apparent fledging success was generally high, complete reproductive failure occurred at one small colony in all years, possibly due to mammalian predators. In 2003, we found a striking difference in diet composition between colonies, with nestlings on Chiniak Island in the outer bay consuming primarily Pacific sand lance Ammodytes hexapterus (50%) and capelin Mallotus villosus (45%) and nestlings at Cliff Island in the inner bay (22 km away) having a diet dominated (76%) by lower quality Pacific sandfish Trichodon trichodon. -

Seasonal Composition and Abundance of Juvenile And
SEASONAL COMPOSITION AND ABUNDANCE OF JUVENILE AND ADULT MARINE FINFISH AND CRAB SPECIES IN THE NEARSHORE ZONE OF KODIAK ISLAND’S EASTSIDE DURING APRIL 1978 THROUGH MARCH 1979 by James E. Blackburn and Peter B. Jackson Alaska Department of Fish and Game Final Report Outer Continental Shelf Environmental Assessment Program Research Unit 552 April 1982 377” TABLE OF CONTENTS E%?!2 List of Figures. 381 List of Tables . 385 List of Appendix Tables. 387 Summary of Objectives and Results with Respect to OCS Oil and Gas Development . 391 Introduction . 392 General Nature and Scope of Study . 392 Specific Objectives . 392 Relevance to Problems of Petroleum Development. 392 Acknowledgements. 392 Current State of Knowledge . 393 King Crab . 394 Tanner Crab . 394 Dungeness Crab. 397 Shrimp. 397 Scallops. 399 Salmon. 399 Herring . 411 Halibut . 412 Bottomfish. 413 Study Area . 413 Sources, Methods and Rationale of Data Collection. 419 Beach Seine . 420 Gill Net. 421 Trammel Net . 421 Tow Net . 421 Try Net . 421 Otter Trawl . 422 Sample Handling . 422 Stages of Maturity. 422 Sample Analysis . 423 Area Comparisons . 424 Diversity. 424 Species Association. 425 Data Limitations. 425 Results. 426 Relative Abundance. 430 Seasonality by Habitat. 437 Nearshore Habitat. 437 Pelagic Habitat. 443 Demersal Habitat . 443 Area Comparisons. 449 Species Associations . 463 Diversity . 465 Features of Distribution, Abundance, Migration, Growth and Reproduction of Prominent Taxa . 473 King Crab.. 473 Tanner Crab. 473 Pacific Herring. 473 Pink Salmon. 473 ChumSalnion. 482 CohoSalmon. 482 DollyVarden. 482 Capelin. 486 Pacific Cod. 486 PacificTomcod. ---- . 489 WalleyePollock. 489 Rockfish. 489 RockGreenling. 493 MaskedGreenling. 493 Whitespotted Greenling. 495 Seblefish. -

RACE Species Codes and Survey Codes 2018
Alaska Fisheries Science Center Resource Assessment and Conservation Engineering MAY 2019 GROUNDFISH SURVEY & SPECIES CODES U.S. Department of Commerce | National Oceanic and Atmospheric Administration | National Marine Fisheries Service SPECIES CODES Resource Assessment and Conservation Engineering Division LIST SPECIES CODE PAGE The Species Code listings given in this manual are the most complete and correct 1 NUMERICAL LISTING 1 copies of the RACE Division’s central Species Code database, as of: May 2019. This OF ALL SPECIES manual replaces all previous Species Code book versions. 2 ALPHABETICAL LISTING 35 OF FISHES The source of these listings is a single Species Code table maintained at the AFSC, Seattle. This source table, started during the 1950’s, now includes approximately 2651 3 ALPHABETICAL LISTING 47 OF INVERTEBRATES marine taxa from Pacific Northwest and Alaskan waters. SPECIES CODE LIMITS OF 4 70 in RACE division surveys. It is not a comprehensive list of all taxa potentially available MAJOR TAXONOMIC The Species Code book is a listing of codes used for fishes and invertebrates identified GROUPS to the surveys nor a hierarchical taxonomic key. It is a linear listing of codes applied GROUNDFISH SURVEY 76 levelsto individual listed under catch otherrecords. codes. Specifically, An individual a code specimen assigned is to only a genus represented or higher once refers by CODES (Appendix) anyto animals one code. identified only to that level. It does not include animals identified to lower The Code listing is periodically reviewed