Spatial and Temporal Variation in Tufted Puffin Fratercula Cirrhata Nestling Diet Quality and Growth Rates
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Tufted Puffin Listing Decision
For Immediate Release State Applauds Decision Not to List Tufted Puffin under the Endangered Species Act Dec. 2, 2020 (Juneau) – The State of Alaska is pleased with the “not warranted” finding issued today by the U.S. Fish and Wildlife Service (USFWS or Service), finding that listing the Tufted Puffin (Fratercula cirrhata) under the Endangered Species Act (ESA) is not justified by the best available science. This finding is the culmination of a range-wide status review of the species, which was triggered by a 2014 petition from the Natural Resources Defense Council. The Tufted Puffin is a colorful, medium-sized seabird that breeds in the coastal U.S. states and Canada from California north to Alaska, as well as in Russia and Japan. Citing threats such as a declining prey base, oil spills, and climate change, the petition focused on apparent declines in the contiguous U.S. or “Lower 48” population. The petitioners requested listing of the entire species if the contiguous U.S. population did not meet the definition of a Distinct Population Segment. The Service found that it did not and proceeded to evaluate listing the species across its range. The majority of Tufted Puffins nest in Alaska. A listing under the ESA would have resulted in regulations that likely would have affected Alaska coastal communities, commercial fisheries, and tourism activities. The Alaska Department of Fish and Game (ADF&G) helped collect, collate, and provide population and other data to the Service and reviewed the Service’s Species Status Assessment, which was the basis for their not warranted decision. -

Fishery Management Plan for Groundfish of the Bering Sea and Aleutian Islands Management Area APPENDICES
FMP for Groundfish of the BSAI Management Area Fishery Management Plan for Groundfish of the Bering Sea and Aleutian Islands Management Area APPENDICES Appendix A History of the Fishery Management Plan ...................................................................... A-1 A.1 Amendments to the FMP ......................................................................................................... A-1 Appendix B Geographical Coordinates of Areas Described in the Fishery Management Plan ..... B-1 B.1 Management Area, Subareas, and Districts ............................................................................. B-1 B.2 Closed Areas ............................................................................................................................ B-2 B.3 PSC Limitation Zones ........................................................................................................... B-18 Appendix C Summary of the American Fisheries Act and Subtitle II ............................................. C-1 C.1 Summary of the American Fisheries Act (AFA) Management Measures ............................... C-1 C.2 Summary of Amendments to AFA in the Coast Guard Authorization Act of 2010 ................ C-2 C.3 American Fisheries Act: Subtitle II Bering Sea Pollock Fishery ............................................ C-4 Appendix D Life History Features and Habitat Requirements of Fishery Management Plan SpeciesD-1 D.1 Walleye pollock (Theragra calcogramma) ............................................................................ -

Tufted Puffin
Interpretative Fact Sheet Tufted Puffin (Fratercula cirrhata) The following short article is from the Oregon Coast 101 Species collection used by the Guide and Outfitter Recognized Professional (GORP) training program. These articles are intended to provide interesting facts you can share with your clientele and add value to your services. An Interpretive Fact Sheet has been written about each species. We are currently uploading these blogs and creating the links. Come visit us! Tourism and Business Development College of Business, Oregon State University Extension - Oregon Sea Grant at http://tourism.oregonstate.edu/ Guide and Outfitter Recognized Professional Program https://www.GORPguide.org For more information about the GORP training program see: https://www.gorpguide.org/become-a-gorp-certified-guide Tufted Puffin (Fratercula cirrhata) tourism.oregonstate.edu/tufted-puffin-fratercula-cirrhata/ By colliek2 August 28, 2020 ‘Small Axe’ The tufted puffin is a familiar bird on the Russian and U.S. Pacific coasts. In Russia it is known as ‘toporok’ meaning “small axe.” Not only is this a nod to the shape of the bill but also to one of the main breeding sites, Kamen Toporkov (“Tufted Puffin Rock”) which is an islet offshore Bering Island. Mature, breeding adult Tufted Puffin (royalty free Unsplash) Local Finds We don’t have to go to Russia to see them as they are also found in our backyards (so to speak). This recognizable seabird nests on Oregon headland such as Cape Mears, Cape Lookout, Cape Foulweather, Yaquina Head, and further north at Three Arch Rocks. 1/4 Recognizing The large triangular red-orange bill is definitely unique and is most visible on breeding adults during the summer reproductive season. -

2020-2021 Statewide Commercial Groundfish Fishing Regulations
Alaska Department of Fish and Game 2020–2021 Statewide Commercial Groundfish Fishing Regulations This booklet contains regulations regarding COMMERCIAL GROUNDFISH FISHERIES in the State of Alaska. This booklet covers the period May 2020 through March 2021 or until a new book is available following the Board of Fisheries meetings. Note to Readers: These statutes and administrative regulations were excerpted from the Alaska Statutes (AS), and the Alaska Administrative Code (AAC) based on the official regulations on file with the Lieutenant Governor. There may be errors or omissions that have not been identified and changes that occurred after this printing. This booklet is intended as an informational guide only. To be certain of the current laws, refer to the official statutes and the AAC. Changes to Regulations in this booklet: The regulations appearing in this booklet may be changed by subsequent board action, emergency regulation, or emergency order at any time. Supplementary changes to the regulations in this booklet will be available on the department′s website and at offices of the Department of Fish and Game. For information or questions regarding regulations, requirements to participate in commercial fishing activities, allowable activities, other regulatory clarifications, or questions on this publication please contact the Regulations Program Coordinator at (907) 465-6124 or email [email protected] The Alaska Department of Fish and Game (ADF&G) administers all programs and activities free from discrimination based on race, color, national origin, age, sex, religion, marital status, pregnancy, parenthood, or disability. The department administers all programs and activities in compliance with Title VI of the Civil Rights Act of 1964, Section 504 of the Rehabilitation Act of 1973, Title II of the Americans with Disabilities Act of 1990, the Age Discrimination Act of 1975, and Title IX of the Education Amendments of 1972. -

Breeding Biology of the Horned Puffin on St. Lawrence Island, Bering Sea, with Zoogeographical Notes on the North Pacific Puffins I
Pacific Science (1973), Vol. 27, No.2, p. 99-119 Printed in Great Britain Breeding Biology of the Horned Puffin on St. Lawrence Island, Bering Sea, with Zoogeographical Notes on the North Pacific Puffins I SPENCER G. SEALY' THE HORNED PUFFIN (Fratercula corniculata) is one of six species ofalcids which regularly nest on Sevuokuk Mountain, 3 km east of Gambell on St. Lawrence Island, Alaska (Fig. 1). During the summers of 1966 and 1967, I conducted on this island a study of the breeding ecology of three of these species, the Parakeet Auklet (Cyc/orrf?ynchuspsittacula), Crested Auklet (Aethia cristatella), and Least Auklet (A. pusilla) (see Sealy, 1968). During these summers some ob servations on the breeding biology of the Horn ed Puffin were obtained and are reported here. The only life history study ofthis species which spans the entire breeding season is that of Swartz (1966) in the Cape Thompson region, Alaska, some 560 km north of St. Lawrence Island (Fig. 2). Numerous studies of the biology of the con generic Common Puffin (Fratercula arctica) of the Atlantic and Arctic oceans are available (e.g., Lockley, 1953; Be1opol'skii, 1957; Uspen ski, 1958; Myrberget, 1959, 1961, 1962; Kartas chew, 1960; Nettleship, 1972; and others) and some of these will be utilized here for compara tive purposes. When available, comparative ob servations on the breeding biology of the other Pacific puffins, the Rhinoceros Auklet (Ceror hinca monocerata), which is actually a puffin (Storer, 1945), and the Tufted Puffin (Lunda cirrhata) will also be included. DISTRIBUTION The breeding distribution of the Horned Puffin has been mapped recently by Udvardy (1963: 105). -

Humboldt Bay Fishes
Humboldt Bay Fishes ><((((º>`·._ .·´¯`·. _ .·´¯`·. ><((((º> ·´¯`·._.·´¯`·.. ><((((º>`·._ .·´¯`·. _ .·´¯`·. ><((((º> Acknowledgements The Humboldt Bay Harbor District would like to offer our sincere thanks and appreciation to the authors and photographers who have allowed us to use their work in this report. Photography and Illustrations We would like to thank the photographers and illustrators who have so graciously donated the use of their images for this publication. Andrey Dolgor Dan Gotshall Polar Research Institute of Marine Sea Challengers, Inc. Fisheries And Oceanography [email protected] [email protected] Michael Lanboeuf Milton Love [email protected] Marine Science Institute [email protected] Stephen Metherell Jacques Moreau [email protected] [email protected] Bernd Ueberschaer Clinton Bauder [email protected] [email protected] Fish descriptions contained in this report are from: Froese, R. and Pauly, D. Editors. 2003 FishBase. Worldwide Web electronic publication. http://www.fishbase.org/ 13 August 2003 Photographer Fish Photographer Bauder, Clinton wolf-eel Gotshall, Daniel W scalyhead sculpin Bauder, Clinton blackeye goby Gotshall, Daniel W speckled sanddab Bauder, Clinton spotted cusk-eel Gotshall, Daniel W. bocaccio Bauder, Clinton tube-snout Gotshall, Daniel W. brown rockfish Gotshall, Daniel W. yellowtail rockfish Flescher, Don american shad Gotshall, Daniel W. dover sole Flescher, Don stripped bass Gotshall, Daniel W. pacific sanddab Gotshall, Daniel W. kelp greenling Garcia-Franco, Mauricio louvar -

SHOREBIRDS (Charadriiformes*) CARE MANUAL *Does Not Include Alcidae
SHOREBIRDS (Charadriiformes*) CARE MANUAL *Does not include Alcidae CREATED BY AZA CHARADRIIFORMES TAXON ADVISORY GROUP IN ASSOCIATION WITH AZA ANIMAL WELFARE COMMITTEE Shorebirds (Charadriiformes) Care Manual Shorebirds (Charadriiformes) Care Manual Published by the Association of Zoos and Aquariums in association with the AZA Animal Welfare Committee Formal Citation: AZA Charadriiformes Taxon Advisory Group. (2014). Shorebirds (Charadriiformes) Care Manual. Silver Spring, MD: Association of Zoos and Aquariums. Original Completion Date: October 2013 Authors and Significant Contributors: Aimee Greenebaum: AZA Charadriiformes TAG Vice Chair, Monterey Bay Aquarium, USA Alex Waier: Milwaukee County Zoo, USA Carol Hendrickson: Birmingham Zoo, USA Cindy Pinger: AZA Charadriiformes TAG Chair, Birmingham Zoo, USA CJ McCarty: Oregon Coast Aquarium, USA Heidi Cline: Alaska SeaLife Center, USA Jamie Ries: Central Park Zoo, USA Joe Barkowski: Sedgwick County Zoo, USA Kim Wanders: Monterey Bay Aquarium, USA Mary Carlson: Charadriiformes Program Advisor, Seattle Aquarium, USA Sara Perry: Seattle Aquarium, USA Sara Crook-Martin: Buttonwood Park Zoo, USA Shana R. Lavin, Ph.D.,Wildlife Nutrition Fellow University of Florida, Dept. of Animal Sciences , Walt Disney World Animal Programs Dr. Stephanie McCain: AZA Charadriiformes TAG Veterinarian Advisor, DVM, Birmingham Zoo, USA Phil King: Assiniboine Park Zoo, Canada Reviewers: Dr. Mike Murray (Monterey Bay Aquarium, USA) John C. Anderson (Seattle Aquarium volunteer) Kristina Neuman (Point Blue Conservation Science) Sarah Saunders (Conservation Biology Graduate Program,University of Minnesota) AZA Staff Editors: Maya Seaman, MS, Animal Care Manual Editing Consultant Candice Dorsey, PhD, Director of Animal Programs Debborah Luke, PhD, Vice President, Conservation & Science Cover Photo Credits: Jeff Pribble Disclaimer: This manual presents a compilation of knowledge provided by recognized animal experts based on the current science, practice, and technology of animal management. -

Tufted Puffin Monitoring Study at Haystack Rock, Cannon Beach, Oregon 2010-2013
Tufted Puffin Monitoring Study at Haystack Rock, Cannon Beach, Oregon 2010-2013 Photo credit USFWS By Shawn W. Stephensen U.S. Fish and Wildlife Service Oregon Coast National Wildlife Refuge Complex 2127 SE Marine Science Drive Newport, Oregon 97365 January 2014 Cite as: Stephensen, S.W. 2014. Tufted Puffin monitoring study at Haystack Rock, Cannon Beach, Oregon 2010-2013. U.S. Fish and Wildlife Service Unpublished Report, Oregon Coast National Wildlife Refuge Complex, Newport, Oregon 97365. 16 pp. EXECUTIVE SUMMARY The Tufted Puffin (Fratercula cirrhata) is a medium-large pelagic seabird and member of the Auk family. The distribution of the Tufted Puffin is widespread in the North Pacific Ocean and nests on the coastline and offshore islands in California, Oregon, Washington, British Columbia, Alaska, Japan, and Russia. Tufted Puffin populations have generally declined throughout the southern portion of their range from British Columbia to northern California. Possible causes of puffin decline include factors related to conditions at breeding sites, at-sea mortality due to direct human impacts, and long-term changes in marine food webs. The U. S. Fish and Wildlife Service conducted a burrow-nesting seabird survey that encompassed the entire coastline of Oregon in 2008 and documented an order of magnitude decline in the puffin population since the previous official statewide survey in 1988. The purpose of this project was to conduct an intensive population status assessment of the Tufted Puffin at Haystack Rock (colony number 219-021), which is part of the Oregon Islands National Wildlife Refuge. Haystack Rock is a 72 meter tall sea stack, located on the north coast of Oregon at Cannon Beach. -

TUFTED PUFFIN Fratercula Cirrhata
Alaska Seabird Information Series TUFTED PUFFIN Fratercula cirrhata Conservation Status ALASKA: Not At Risk N. AMERICAN: Moderate Concern GLOBAL: Least Concern Breed Eggs Incubation Fledge Nest Feeding Behavior Diet May-Sept 1 40-53 d 45-55 d burrow, crevice surface dive fish, squid, other invertebrates Life History and Distribution Tufted puffins (Fratercula cirrhata) are highly decorative seabirds. Breeding adults have huge orange bills, legs, and feet, white faces, and long golden feather tufts that curl back from each side of the head. In late summer, they lose their tufts and the bright colors of the bill turn to a dull reddish-brown. Diet is one of the fascinating details of Tufted Puffin st ur biology. Chicks are fed almost entirely tiny fish which the wh parents catch underwater and collect, lined up head to tail, De nna across their bills. They routinely hold 5-20 fish in their o mouths while returning to the nest. Puffins use their S D tongues to hold the fish against the spiny palate in their mouths while opening their bill to catch more fish. USFW This species prefers high, steep areas for nesting. Although they are about the size of a crow, they are twice Alaska Seasonal Distribution as heavy with short, stubby wings. The wings are used for AK Region Sp S F W “flying” underwater in pursuit of food; this same feature Southeastern * U U U R makes them poor aerial flyers. Tall cliffs make for easy Southcoastal * C C C R take-offs and give newly fledged puffins assistance in Southwestern * C C C U getting up enough airspeed for their first flight. -

Page 1 . ., = C ===(/) ···===, V'f---~ =(F) ~-:R: '10-"' Oo===E N =~ = R =A
., ===(/)=c ···====,V'f---~=(f) ~-:r: /lc)L;J '10-"' oo===E /3?/ N =~ =r==r=a o=~==, 0 =[11 o===oo N==m oo===~ N== ..... =n ~==[11 Census of Seabirds Nesting on The Triplets Islands, June 1985 Vl =])=I=' oo::==::::])=r =,..=oo -]) 'LIBRARY USFWS BY Anchorage Catherine M. Berg* Douglas J. Forse11t Kenneth R. Emmel* Key Words: Kodiak Island, The Triplets Islands, Abundance, Tufted Puffin, Fork-tailed Storm-Petrel, Leach's Storm-Petrel, Ancient Murrelet, Gulls, Subsistence * Division of Realty t Alaska Office of Fish and Wildlife Research U.S. Fish and Wildlife Service 1011 E. Tudor Road Anchorage, Alaska 99503 July 1985 INTRODUCTION The Triplets Islands is the largest seabird rookery in the Kodiak archipelago (Figure 1). These islands were surveyed by boat in 1975 by Matt Dick and in 1977 by John Trapp, Matt Dick, Doug Forsell, and Eric Holberg. We surveyed the Triplet Islands in late June, 1985, accompanied by Richard Macintosh and Tony DeGange. This was the first survey conducted on land and the first nocturnal surveys. The purpose of this trip was to accurately assess the seabird resources of The Triplets and determine their value for acquisition by the U.S. Fish and Wildlife Service. This acquisition would be part of a land exchange proposal at Ugashik Narrows on the Alaska Peninsula NWR. Table 1. Itinerary of The Triplets Islands Survey, 1985. DATE ACTIVITY June 21 Doug Forsell, Cathy Berg, Ken Emmel, and Rich Macintosh departed Kodiak, for The Triplets, aboard a 13-foot Zodiak and a 16-foot whaler. Route followed Woody I. Channel, northwest around Spruce Cape, through Narrow Strait, and then north to The Triplets. -

Guide to the Coastal Marine Fishes of California
STATE OF CALIFORNIA THE RESOURCES AGENCY DEPARTMENT OF FISH AND GAME FISH BULLETIN 157 GUIDE TO THE COASTAL MARINE FISHES OF CALIFORNIA by DANIEL J. MILLER and ROBERT N. LEA Marine Resources Region 1972 ABSTRACT This is a comprehensive identification guide encompassing all shallow marine fishes within California waters. Geographic range limits, maximum size, depth range, a brief color description, and some meristic counts including, if available: fin ray counts, lateral line pores, lateral line scales, gill rakers, and vertebrae are given. Body proportions and shapes are used in the keys and a state- ment concerning the rarity or commonness in California is given for each species. In all, 554 species are described. Three of these have not been re- corded or confirmed as occurring in California waters but are included since they are apt to appear. The remainder have been recorded as occurring in an area between the Mexican and Oregon borders and offshore to at least 50 miles. Five of California species as yet have not been named or described, and ichthyologists studying these new forms have given information on identification to enable inclusion here. A dichotomous key to 144 families includes an outline figure of a repre- sentative for all but two families. Keys are presented for all larger families, and diagnostic features are pointed out on most of the figures. Illustrations are presented for all but eight species. Of the 554 species, 439 are found primarily in depths less than 400 ft., 48 are meso- or bathypelagic species, and 67 are deepwater bottom dwelling forms rarely taken in less than 400 ft. -
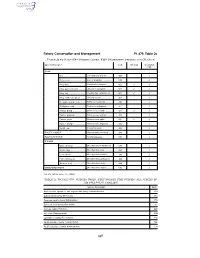
Species Codes: Fmp Prohibited Species and Cr Crab
Fishery Conservation and Management Pt. 679, Table 2c TABLE 2b TO PART 679—SPECIES CODES: FMP PROHIBITED SPECIES AND CR CRAB Species Description Code CR Crab Groundfish PSC CRAB Box ....................................... Lopholithodes mandtii .......... 900 ✓ Dungeness ........................... Cancer magister .................. 910 ✓ King, blue ............................. Paralithodes platypus .......... 922 ✓ ✓ King, golden (brown) ........... Lithodes aequispinus ........... 923 ✓ ✓ King, red .............................. Paralithodes camtshaticus ... 921 ✓ ✓ King, scarlet (deepsea) ....... Lithodes couesi .................... 924 ✓ Korean horsehair crab ......... Erimacrus isenbeckii ............ 940 ✓ Multispinus crab ................... Paralomis multispinus .......... 951 ✓ Tanner, Bairdi ...................... Chionoecetes bairdi ............. 931 ✓ ✓ Tanner, grooved .................. Chionoecetes tanneri ........... 933 ✓ Tanner, snow ....................... Chionoecetes opilio ............. 932 ✓ ✓ Tanner, triangle ................... Chionoecetes angulatus ...... 934 ✓ Verrilli crab ........................... Paralomis verrilli .................. 953 ✓ PACIFIC HALIBUT Hippoglossus stenolepis ...... 200 ✓ PACIFIC HERRING Family Clupeidae ................. 235 ✓ SALMON Chinook (king) ..................... Oncorhynchus tshawytscha 410 ✓ Chum (dog) .......................... Oncorhynchus keta .............. 450 ✓ Coho (silver) ........................ Oncorhynchus kisutch ......... 430 ✓ Pink (humpback) .................