Interspecific Lily Hybrids with the Ability to Flower Precociously and to Produce Multiple Flower Stalks from Lilium Formosanum
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Guide to the Flora of the Carolinas, Virginia, and Georgia, Working Draft of 17 March 2004 -- LILIACEAE
Guide to the Flora of the Carolinas, Virginia, and Georgia, Working Draft of 17 March 2004 -- LILIACEAE LILIACEAE de Jussieu 1789 (Lily Family) (also see AGAVACEAE, ALLIACEAE, ALSTROEMERIACEAE, AMARYLLIDACEAE, ASPARAGACEAE, COLCHICACEAE, HEMEROCALLIDACEAE, HOSTACEAE, HYACINTHACEAE, HYPOXIDACEAE, MELANTHIACEAE, NARTHECIACEAE, RUSCACEAE, SMILACACEAE, THEMIDACEAE, TOFIELDIACEAE) As here interpreted narrowly, the Liliaceae constitutes about 11 genera and 550 species, of the Northern Hemisphere. There has been much recent investigation and re-interpretation of evidence regarding the upper-level taxonomy of the Liliales, with strong suggestions that the broad Liliaceae recognized by Cronquist (1981) is artificial and polyphyletic. Cronquist (1993) himself concurs, at least to a degree: "we still await a comprehensive reorganization of the lilies into several families more comparable to other recognized families of angiosperms." Dahlgren & Clifford (1982) and Dahlgren, Clifford, & Yeo (1985) synthesized an early phase in the modern revolution of monocot taxonomy. Since then, additional research, especially molecular (Duvall et al. 1993, Chase et al. 1993, Bogler & Simpson 1995, and many others), has strongly validated the general lines (and many details) of Dahlgren's arrangement. The most recent synthesis (Kubitzki 1998a) is followed as the basis for familial and generic taxonomy of the lilies and their relatives (see summary below). References: Angiosperm Phylogeny Group (1998, 2003); Tamura in Kubitzki (1998a). Our “liliaceous” genera (members of orders placed in the Lilianae) are therefore divided as shown below, largely following Kubitzki (1998a) and some more recent molecular analyses. ALISMATALES TOFIELDIACEAE: Pleea, Tofieldia. LILIALES ALSTROEMERIACEAE: Alstroemeria COLCHICACEAE: Colchicum, Uvularia. LILIACEAE: Clintonia, Erythronium, Lilium, Medeola, Prosartes, Streptopus, Tricyrtis, Tulipa. MELANTHIACEAE: Amianthium, Anticlea, Chamaelirium, Helonias, Melanthium, Schoenocaulon, Stenanthium, Veratrum, Toxicoscordion, Trillium, Xerophyllum, Zigadenus. -

Historical Survey of Easter Lily Name in Association with Lilium Longiflorum
® Floriculture and Ornamental Biotechnology ©2012 Global Science Books Historical Survey of Easter Lily Name in Association with Lilium longiflorum Eisuke Matsuo Professor Emeritus of Kyushu University, 1198-68, Tsuko, Ogori-shi 838-0102, Japan Correspondence : [email protected] ABSTRACT Based on the English dictionaries from around 1900, the author traces back how Lilium longiflorum came to be referred to as the Easter lily. The term “Easter lily” was given to any lily-like flowering species that bloomed around the Easter day, being called Easter flowers. Among them, L. candidum became the most famous one known as “Easter lily”. The evidence suggests that when more easily forcible bulb production began in the Bermuda Islands, the superiority of L. longiflorum over L. candidum in forcing ability and price, might have led to an explosive promotion of the usage of L. longiflorum as the Easter flower instead of L. candidum. Thus, the name "Easter lily" came to be transferred from L. candidum to L. longiflorum. _____________________________________________________________________________________________________________ Keywords: Easter decoration, Easter flower, Lilium candidum, Madonna lily CONTENTS INTRODUCTION.......................................................................................................................................................................................... 9 EASTER FLOWER AND EASTER LILY................................................................................................................................................... -

Overexpression of Lilium Formosanum MADS-Box (LFMADS) Causing
International Journal of Molecular Sciences Article Overexpression of Lilium formosanum MADS-box (LFMADS) Causing Floral Defects While Promoting Flowering in Arabidopsis thaliana, Whereas Only Affecting Floral Transition Time in Nicotiana tabacum Wan-Yu Liao 1,†, Lee-Fong Lin 2,† ID , Ming-Der Lin 3, Sheng-Che Hsieh 2, Althea Yi-Shan Li 2, Yueh-Shiah Tsay 4 and Ming-Lun Chou 1,2,* ID 1 Institute of Medical Sciences, Tzu-Chi University, Hualien 97004, Taiwan; [email protected] 2 Department of Life Sciences, Tzu-Chi University, Hualien 97004, Taiwan; [email protected] (L.-F.L.); [email protected] (S.-C.H.); [email protected] (A.Y.-S.L.) 3 Department of Molecular Biology and Human Genetics, Tzu-Chi University, Hualien 97004, Taiwan; [email protected] 4 Division of Crop Improvement, Hualien District Agricultural Research and Extension Station, Council of Agriculture, Executive Yuan, Hualien 97365, Taiwan; [email protected] * Correspondence: [email protected]; Tel.: +886-3-8465615; Fax: +886-3-8572526 † These authors contributed equally to this study. Received: 30 June 2018; Accepted: 26 July 2018; Published: 29 July 2018 Abstract: The Formosa lily (Lilium formosanum) is one of the most common horticultural species in Taiwan. To explore gene regulation involved in this species, we used transcriptome analysis to generate PH-FB (mixed floral buds) and PH-LF (mature leaves) datasets. Combination of the PH-FB and PH-LF constructed a de novo assembly of the ALL dataset, including 18,041 contigs and 23,807 unigenes by Nr, GO, COG, and KEGG databases. -
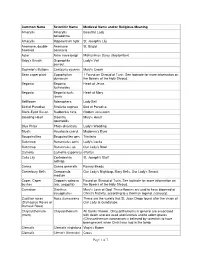
Of 7 Common Name Scientific Name Medieval Name And/Or Religious Meaning Amaryllis Amaryllis Belladonna Beautiful Lady
Common Name Scientific Name Medieval Name and/or Religious Meaning Amaryllis Amaryllis Beautiful Lady belladonna Amaryllis Hippeastrum hybr. St. Joseph's Lily Anemone, double- Anemone St. Brigid flowered coronaria Aster Aster nova-belgii Michaelmas Daisy (September) Baby's Breath Gypsophila Lady's Veil panicul. Bachelor's Buttons Centauria cyannis Mary's Crown Bean caper plant Zygophyllum ? Found on Shroud of Turin. See footnote for more information on dumosum the flowers of the Holy Shroud. Begonia Begonia Heart of Jesus fuchsioides Begonia Begonia fuch. Heart of Mary rosea Bellflower Adenophera Lady Bell Bird of Paradise Streliztia reginae Bird of Paradise Black-Eyed Susan Rudbeckia hirta Golden Jerusalem Bleeding Heart Dicentra Mary's Heart spectabilis Blue Phlox Phlox divaricata Lady's Wedding Bluets Houstonia caerul. Madonna's Eyes Bougainvillea Bougainvillea gen. Trinitaria Buttercup Ranunculus acris Lady's Locks Buttercup Ranunculus sp. Our Lady's Bowl Camelia Camellia (japonica) (Purity) Calla Lily Zantedeshia St. Joseph's Staff aethiop. Canna Canna generalis Rosary Beads Canterbury Bells Campanula Our Lady's Nightcap, Mary Bells, Our Lady's Smock medium Caper, Caper Capparis spinosa Found on Shroud of Turin. See footnote for more information on bushes (var. aegyptia) the flowers of the Holy Shroud. Carnation Dianthus Mary's Love of God. These flowers are said to have bloomed at caryophyllus Christ's Nativity, according to a German legend. (January) Castilian roses Rosa damascena These are the variety that St. Juan Diego found after the vision of (Damascus Roses or Our Lady at Guadalupe. Damask Rose) Chrysanthemum Chrysanthemum All Saints' Flower. Chrysanthemums in general are associated (mum) with death and are used and funerals and to adorn graves (Chrysanthemum coronarium is believed by scientists to have been present when Christ was laid in the tomb. -

LILIUM) PRODUCTION Faculty of Science, Department of Biology, University of Oulu
BIOTECHNOLOGICAL APPROACHES VELI-PEKKA PELKONEN IN LILY (LILIUM) PRODUCTION Faculty of Science, Department of Biology, University of Oulu OULU 2005 VELI-PEKKA PELKONEN BIOTECHNOLOGICAL APPROACHES IN LILY (LILIUM) PRODUCTION Academic Dissertation to be presented with the assent of the Faculty of Science, University of Oulu, for public discussion in Kuusamonsali (Auditorium YB210), Linnanmaa, on April 15th, 2005, at 12 noon OULUN YLIOPISTO, OULU 2005 Copyright © 2005 University of Oulu, 2005 Supervised by Professor Anja Hohtola Professor Hely Häggman Reviewed by Professor Anna Bach Professor Risto Tahvonen ISBN 951-42-7658-2 (nid.) ISBN 951-42-7659-0 (PDF) http://herkules.oulu.fi/isbn9514276590/ ISSN 0355-3191 http://herkules.oulu.fi/issn03553191/ OULU UNIVERSITY PRESS OULU 2005 Pelkonen, Veli-Pekka, Biotechnological approaches in lily (Lilium) production Faculty of Science, Department of Biology, University of Oulu, P.O.Box 3000, FIN-90014 University of Oulu, Finland 2005 Oulu, Finland Abstract Biotechnology has become a necessity, not only in research, but also in the culture and breeding of lilies. Various methods in tissue culture and molecular breeding have been applied to the production of commercially important lily species and cultivars. However, scientific research data of such species and varieties that have potential in the northern climate is scarce. In this work, different biotechnological methods were developed and used in the production and culture of a diversity of lily species belonging to different taxonomic groups. The aim was to test and develop further the existing methods in plant biotechnology for the developmental work and the production of novel hardy lily cultivars for northern climates. -

Easter Lily Growth and Development
Easter Lily Growth and Development 1985 Lot aClef - uNIPR TqTE .` cam V /rr sl 81. ,! l9' Technical Bulletin 148 AGRICULTURAL EXPERIMENT STATION Oregon State University Corvallis, Oregon Easter Lily Growth and Development ACKNOWLEDGMENTS We wish to acknowledge the help and encouragement of colleagues concernedwith lily bulb growing, handling, and forcing at Oregon State over the years. Faculty members including L. T.Blaney,O. C.Compton. Ralph Garren, L. H.Fuchigami, J. C.Green, P. J. Breen, and J. R. Stang have made important contributions to our knowledge of Lilium physiology. Graduate students in the Department of Horticulture have contributed much of thedetail to thisknowledge. These graduates include Vernon Clarkson (M.S. 1951),P. C. Lin(M.S. 1968). Dave Hartley (Ph.D.1968). S. Y. Wang(Ph.D.1969).Raj Bahadur(Ph.D. 1969), W. R. McCorkle(B.S. 1970, Missouri),K. D. Gray(M.S. 1974), B. J. Tomasovic(B.S.1974),and Y. T. Wang(M.S. 1981). We are especially grateful tothe FredC. Cloeckncr Foundation for its many years of financial support of the Easter lily research project and in preparation of this publication. The same is true for the Pacific Bulb Growers' Association, which has provided a field research station, a technician for its maintenance, graduate student support, and operating expenses at the station for more than 25 years. We would also mention the valuable contributions made by the Agricultural Extension Services of both Oregon and California through their able representatives Walt Schroeder (Curry County, Oregon) and John Lenz (Del Norte County, California). We are indebted to H. -

Evolutionary Events in Lilium (Including Nomocharis, Liliaceae
Molecular Phylogenetics and Evolution 68 (2013) 443–460 Contents lists available at SciVerse ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Evolutionary events in Lilium (including Nomocharis, Liliaceae) are temporally correlated with orogenies of the Q–T plateau and the Hengduan Mountains ⇑ Yun-Dong Gao a,b, AJ Harris c, Song-Dong Zhou a, Xing-Jin He a, a Key Laboratory of Bio-Resources and Eco-Environment of Ministry of Education, College of Life Science, Sichuan University, Chengdu 610065, China b Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China c Department of Botany, Oklahoma State University, Oklahoma 74078-3013, USA article info abstract Article history: The Hengduan Mountains (H-D Mountains) in China flank the eastern edge of the Qinghai–Tibet Plateau Received 21 July 2012 (Q–T Plateau) and are a center of great temperate plant diversity. The geological history and complex Revised 24 April 2013 topography of these mountains may have prompted the in situ evolution of many diverse and narrowly Accepted 26 April 2013 endemic species. Despite the importance of the H-D Mountains to biodiversity, many uncertainties Available online 9 May 2013 remain regarding the timing and tempo of their uplift. One hypothesis is that the Q–T Plateau underwent a final, rapid phase of uplift 8–7 million years ago (Mya) and that the H-D Mountains orogeny was a sep- Keywords: arate event occurring 4–3 Mya. To evaluate this hypothesis, we performed phylogenetic, biogeographic, Hengduan Mountains divergence time dating, and diversification rate analyses of the horticulturally important genus Lilium, Lilium–Nomocharis complex Intercontinental dispersal including Nomocharis. -

20. LILIUM Linnaeus, Sp. Pl. 1: 302. 1753. 百合属 Bai He Shu Liang Songyun (梁松筠 Liang Song-Jun); Minoru N
Flora of China 24: 135–149. 2000. 20. LILIUM Linnaeus, Sp. Pl. 1: 302. 1753. 百合属 bai he shu Liang Songyun (梁松筠 Liang Song-jun); Minoru N. Tamura Herbs perennial, bulbiferous. Bulb of many imbricate, fleshy scales, without tunic. Stem erect, leafy. Leaves alternate, rarely whorled, sessile or subsessile, usually linear to linear-lanceolate. Bulblets sometimes present in leaf axils. Inflorescence terminal, a raceme or solitary flower, very rarely an umbel or corymb; bracts leaflike. Flowers often funnelform or campanulate, sometimes tub- ular or cupular. Tepals 6, free, usually connivent, sometimes strongly recurved or revolute, white, yellow, greenish, or reddish to pur- plish, nectariferous near base adaxially; nectaries usually narrowly grooved, sometimes fringed with papillae or hairs, rarely flat on outer tepals. Stamens 6; filaments subulate or filiform, sometimes pubescent; anthers dorsifixed, versatile. Ovary 3-loculed; ovules many per locule. Style elongate, slender; stigma swollen, usually 3-lobed. Fruit a loculicidal capsule. Seeds many, arranged like a pile of coins in each valve, flat, narrowly winged all round. About 115 species: temperate and alpine regions of the N hemisphere, especially in E Asia; 55 species (35 endemic, one introduced) in China. The status of Lilium puerense Y. Y. Qian (Guihaia 11: 125. 1991) and L. rockii R. H. Miao (Acta Scient. Nat. Univ. Sunyatseni 34(3): 81. 1995) is unclear. Lilium puerense was described from S Yunnan (Pu’er Xian), based on specimens collected in 1987 (holotype: Y. Y. Qian 1774, SMAO). It is said to be similar to L. sulphureum, but with leaf margin papillose, bracts ovate, and ovary greenish (vs. -

60 Uiz R. Prameela
UIZ Q R. PRAMEELA UN F 1. It gives sweetly scented 5. It is called ‘Blood lily’, fl owers and it also called blooms from May to July ‘Amazon lily’, blooms and it has been used as from November to a component of arrow January. poison and fi shing poison. a. Eucharis grandifl ora a. Hippeastrum b. Hymenocallis li oralis b. Amaryllis belladonna c. Zephyranthes grandifl ora c. Scadoxus mul fl orus d. Pancra um trifl orum d. Zephyranthes rosea 6. It means beau ful membrane, blooms in rainy season with white 2. This lily is na ve of fragrant fl owers. It is also South Africa, having long called ‘Spider lily’. leaves. It is also called ‘belladonna lily’ or ‘naked a. Proiphys alba lady’. It contains alkaloid b. Hymenocallis li oralis bellarmine. c. Eucharis grandifl ora a. Scadoxus mul fl orus d. Pancra um trifl orum b. Atropa belladonna c. Amaryllis belladonna d. Crinum asia cum 7. Zephyranthes species are collec vely called rain lilies or Fairy lilies, Zephyr-Flower or Thunder-Flower. They are ny fl owering plants ideal for edgings to borders. They are available in White, Pink, and Yellow. Which of the following is ‘White Zephyr’, blossoms in sunny days? 3. It is a popular indoor ornamental plant; commonly known as ‘Knight’s-star-lily’. a. Hippeastrum b. Lilium c. Crinum a. Zephyranthes candida b. Z. rosea d. Amaryllis c. Z. aurea d. Z. carinata 8. Iden fy the ‘pink Zephyr- 4. Leaves broadly ovate Flower’, it blossoms from to round and leathery. July to December. -

Regional Landscape Surveillance for New Weed Threats Project 2018-2019
State Herbarium of South Australia Botanic Gardens and State Herbarium Economic & Sustainable Development Group Department for Environment and Water Milestone Report Regional Landscape Surveillance for New Weed Threats Project 2018-2019 Milestone: Annual report on new plant naturalisations in South Australia Chris J. Brodie, Peter J. Lang & Michelle Waycott June 2019 Contents Summary............................................................................................................................... 3 1. Activities and outcomes for 2017/2018 financial year........................................................ 3 Funding ............................................................................................................................. 3 Activities ........................................................................................................................... 4 Outcomes and progress of weeds monitoring ..................................................................... 6 2. New naturalised or questionably naturalised records of plants in South Australia. ............. 7 3. Descriptions of newly recognised weeds in South Australia .............................................. 9 4. Updates to weed distributions in South Australia, weed status and name changes ............ 29 References .......................................................................................................................... 33 Appendix 1: Activities of the Weeds Botanist .................................................................... -

The Root Systems of Onion and Allium Fistulosum in The
Flower development of Lilium longiflorum: Characterisation of MADS-box transcription factors VAGNER AUGUSTO BENEDITO Promotor: Prof. dr. S. C. de Vries Hoogleraar in de Biochemie Wageningen Universiteit Co-promotoren: Dr. F. A. Krens Clusterleider, businessunit Biodiversiteit en Veredeling, Plant Research International Prof. dr. Ir. G. C. Angenent Clusterleider, businessunit Bioscience, Plant Research International / Hoogleraar in de Moleculaire Plantontwikkeling, Katholieke Universiteit Nijmegen Promotiecommissie: Dr. L. Colombo Università delgi Studi di Milano, Italië Prof. dr. T. Bisseling Wageningen Universiteit Dr. R. G. H. Immink Plant Research International Prof. dr. L. H. W. van der Plas Wageningen Universiteit Flower development of Lilium longiflorum: Characterisation of MADS-box transcription factors Vagner Augusto Benedito Proefschrift ter verkrijging van de graad van doctor op gezag van de rector magnificus van Wageningen Universiteit Prof. dr. ir. L. Speelman in het openbaar te verdedigen op woensdag 14 januari 2004 des namiddags te vier uur in de Aula Flower development of Lilium longiflorum: Characterisation of MADS-box transcription factors. Ph.D. thesis, Wageningen University, Wageningen, The Netherlands 2004. With summaries in English, Dutch and Portuguese. ISBN: 90-5808-947-9 "To the people from Brazil, I dedicate this thesis" Por isso na impaciência Desta sede de saber, Como as aves do deserto As almas buscam beber… Oh! Bendito o que semeia Livros… livros à mão cheia… E manda o povo pensar! O livro caindo n'alma É germe -

Final Policy Review: Alternative Risk Management Measures to Import Lilium Spp
International plant protection convention 14_EWGCutFlowers_2014_June Final policy review Lilium spp. Agenda Item: 4.1 ------------------------------------------------------------------------------------------------------------------------------------ ------------------------------------------------------------------------------------------------- Final policy review: Alternative risk management measures to import Lilium spp. cut flowers from Taiwan December 2013 International plant protection convention 14_EWGCutFlowers_2014_June Final policy review Lilium spp. Agenda Item: 4.1 ------------------------------------------------------------------------------------------------------------------------------------ ------------------------------------------------------------------------------------------------- © Commonwealth of Australia Ownership of intellectual property rights Unless otherwise noted, copyright (and any other intellectual property rights, if any) in this publication is owned by the Commonwealth of Australia (referred to as the Commonwealth). Creative Commons licence All material in this publication is licensed under a Creative Commons Attribution 3.0 Australia Licence, except for content supplied by third parties, photographic images, logos, and the Commonwealth Coat of Arms. Creative Commons Attribution 3.0 Australia Licence is a standard form licence agreement that allows you to copy, distribute, transmit and adapt this publication provided that you attribute the work. A summary of the licence terms is available from creativecommons.org/licenses/by/3.0/au/deed.en.