Antagonism of the Prostaglandin D Receptors DP and CRTH2 As An
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

CXCL13/CXCR5 Interaction Facilitates VCAM-1-Dependent Migration in Human Osteosarcoma
International Journal of Molecular Sciences Article CXCL13/CXCR5 Interaction Facilitates VCAM-1-Dependent Migration in Human Osteosarcoma 1, 2,3,4, 5 6 7 Ju-Fang Liu y, Chiang-Wen Lee y, Chih-Yang Lin , Chia-Chia Chao , Tsung-Ming Chang , Chien-Kuo Han 8, Yuan-Li Huang 8, Yi-Chin Fong 9,10,* and Chih-Hsin Tang 8,11,12,* 1 School of Oral Hygiene, College of Oral Medicine, Taipei Medical University, Taipei City 11031, Taiwan; [email protected] 2 Department of Orthopaedic Surgery, Chang Gung Memorial Hospital, Puzi City, Chiayi County 61363, Taiwan; [email protected] 3 Department of Nursing, Division of Basic Medical Sciences, and Chronic Diseases and Health Promotion Research Center, Chang Gung University of Science and Technology, Puzi City, Chiayi County 61363, Taiwan 4 Research Center for Industry of Human Ecology and Research Center for Chinese Herbal Medicine, Chang Gung University of Science and Technology, Guishan Dist., Taoyuan City 33303, Taiwan 5 School of Medicine, China Medical University, Taichung 40402, Taiwan; [email protected] 6 Department of Respiratory Therapy, Fu Jen Catholic University, New Taipei City 24205, Taiwan; [email protected] 7 School of Medicine, Institute of Physiology, National Yang-Ming University, Taipei City 11221, Taiwan; [email protected] 8 Department of Biotechnology, College of Health Science, Asia University, Taichung 40402, Taiwan; [email protected] (C.-K.H.); [email protected] (Y.-L.H.) 9 Department of Sports Medicine, College of Health Care, China Medical University, Taichung 40402, Taiwan 10 Department of Orthopedic Surgery, China Medical University Beigang Hospital, Yunlin 65152, Taiwan 11 Department of Pharmacology, School of Medicine, China Medical University, Taichung 40402, Taiwan 12 Chinese Medicine Research Center, China Medical University, Taichung 40402, Taiwan * Correspondence: [email protected] (Y.-C.F.); [email protected] (C.-H.T.); Tel.: +886-4-2205-2121-7726 (C.-H.T.); Fax: +886-4-2233-3641 (C.-H.T.) These authors contributed equally to this work. -

Endogenous Biosynthesis of Prostacyclin and Thromboxane and Platelet Function During Chronic Administration of Aspirin in Man
Endogenous biosynthesis of prostacyclin and thromboxane and platelet function during chronic administration of aspirin in man. G A FitzGerald, … , J A Lawson, A R Brash J Clin Invest. 1983;71(3):676-688. https://doi.org/10.1172/JCI110814. Research Article To assess the pharmacologic effects of aspirin on endogenous prostacyclin and thromboxane biosynthesis, 2,3-dinor-6- keto PGF1 alpha (PGI-M) and 2,3-dinor-thromboxane B2 (Tx-M) were measured in urine by mass spectrometry during continuing administration of aspirin. To define the relationship of aspirin intake to endogenous prostacyclin biosynthesis, sequential urines were initially collected in individuals prior to, during, and subsequent to administration of aspirin. Despite inter- and intra-individual variations, PGI-M excretion was significantly reduced by aspirin. However, full mass spectral identification confirmed continuing prostacyclin biosynthesis during aspirin therapy. Recovery of prostacyclin biosynthesis was incomplete 5 d after drug administration was discontinued. To relate aspirin intake to indices of thromboxane biosynthesis and platelet function, volunteers received 20 mg aspirin daily followed by 2,600 mg aspirin daily, each dose for 7 d in sequential weeks. Increasing aspirin dosage inhibited Tx-M excretion from 70 to 98% of pretreatment control values; platelet TxB2 formation from 4.9 to 0.5% and further inhibited platelet function. An extended study was performed to relate aspirin intake to both thromboxane and prostacyclin generation over a wide range of doses. Aspirin, in the range of 20 to 325 mg/d, resulted in a dose-dependent decline in both Tx-M and PGI-M excretion. At doses of 325-2,600 mg/d Tx-M excretion ranged from 5 to 3% of control values while PGI-M remained at 37-23% of control. -

Challenging the FDA Black Box Warning for High Aspirin Dose with Ticagrelor in Patients with Diabetes James J
PERSPECTIVES IN DIABETES Challenging the FDA Black Box Warning for High Aspirin Dose With Ticagrelor in Patients With Diabetes James J. DiNicolantonio and Victor L. Serebruany – Ticagrelor, a novel reversible antiplatelet agent, has a Food and ASA 300 325 mg (53.6%) in the U.S. compared with the Drug Administration (FDA) black box warning to avoid mainte- rest of the world (1.7%) was the only factor to explain nance doses of aspirin (ASA) .100 mg/daily. This restriction is (out of 37 variables explored) the regional interaction based on the hypothesis that ASA doses .100 mg somehow de- that ticagrelor was less effective and potentially more creased ticagrelor’s benefit in the Platelet Inhibition and Patient harmful than clopidogrel (2). However, this interaction Outcomes (PLATO) U.S. cohort. However, these data are highly was not significant, is highly postrandomized, comes postrandomized, come from a very small subgroup in PLATO from a very small subgroup of PLATO, and makes no (57% of patients in the U.S. site), and make no biological sense. biological sense (3). Moreover, the ticagrelor-ASA interaction was not significant by any multivariate Cox regression analyses. The Complete Re- sponse Review for ticagrelor indicates that for U.S. PLATO patients, an ASA dose .300 mg was not a significant interaction TICAGRELOR-ASA HYPOTHESIS for vascular outcomes. In the ticagrelor-ASA .300 mg cohort, all- The claimed ticagrelor-ASA interaction states that ticagrelor cause and vascular mortality were not significantly increased fi – P plus low-dose ASA is bene cial, whereas increasing doses (hazard ratio [HR] 1.27 [95% CI 0.84 1.93], = 0.262 and 1.39 of ASA in combination with ticagrelor produces adverse [0.87–2.2], P = 0.170), respectively. -

The Role of Inflammatory Pathways in Neuroblastoma Tumorigenesis — Igor Snapkov a Dissertation for the Degree of Philosophiae Doctor – August 2016 Contents
Faculty of Health Sciences Department of Medical Biology Molecular Inflammation Research Group The role of inflammatory pathways in neuroblastoma tumorigenesis — Igor Snapkov A dissertation for the degree of Philosophiae Doctor – August 2016 Contents 1. List of publications .................................................................................................................. 2 2. List of abbreviations ................................................................................................................ 3 3. Introduction ............................................................................................................................. 5 3.1 Cancer and inflammation .................................................................................................. 5 3.2.1 Pattern recognition receptors and danger signals ............................................................ 7 3.2.2 Formyl peptide receptor 1 (FPR1) ................................................................................. 9 3.3.1 Chemokines................................................................................................................. 12 3.3.2 Chemerin ..................................................................................................................... 13 3.4 Neuroblastoma ................................................................................................................ 14 3.5 Hepatic clearance of danger signals ................................................................................ -

GTH 2021 State of the Art—Cardiac Surgery: the Perioperative Management of Heparin-Induced Thrombocytopenia in Cardiac Surgery
Review Article 59 GTH 2021 State of the Art—Cardiac Surgery: The Perioperative Management of Heparin-Induced Thrombocytopenia in Cardiac Surgery Laura Ranta1 Emmanuelle Scala1 1 Department of Anesthesiology, Cardiothoracic and Vascular Address for correspondence Emmanuelle Scala, MD, Centre Anesthesia, Lausanne University Hospital (CHUV), Lausanne, Hospitalier Universitaire Vaudois, Rue du Bugnon 46, BH 05/300, 1011 Switzerland Lausanne, Suisse, Switzerland (e-mail: [email protected]). Hämostaseologie 2021;41:59–62. Abstract Heparin-induced thrombocytopenia (HIT) is a severe, immune-mediated, adverse drug Keywords reaction that paradoxically induces a prothrombotic state. Particularly in the setting of ► Heparin-induced cardiac surgery, where full anticoagulation is required during cardiopulmonary bypass, thrombocytopenia the management of HIT can be highly challenging, and requires a multidisciplinary ► cardiac surgery approach. In this short review, the different perioperative strategies to run cardiopul- ► state of the art monary bypass will be summarized. Introduction genicity of the antibodies and is diagnostic for HIT. The administration of heparin to a patient with circulating Heparin-induced thrombocytopenia (HIT) is a severe, im- pathogenic HITabs puts the patient at immediate risk of mune-mediated, adverse drug reaction that paradoxically severe thrombotic complications. induces a prothrombotic state.1,2 Particularly in the setting The time course of HIT can be divided into four distinct of cardiac surgery, where full anticoagulation is required phases.6 Acute HIT is characterized by thrombocytopenia during cardiopulmonary bypass (CPB), the management of and/or thrombosis, the presence of HITabs, and confirma- HIT can be highly challenging, and requires a multidisciplin- tion of their platelet activating capacity by a functional ary approach. -

Inverse Agonism of SQ 29,548 and Ramatroban on Thromboxane A2 Receptor
Inverse Agonism of SQ 29,548 and Ramatroban on Thromboxane A2 Receptor Raja Chakraborty1,3, Rajinder P. Bhullar1, Shyamala Dakshinamurti2,3, John Hwa4, Prashen Chelikani1,2,3* 1 Department of Oral Biology, University of Manitoba, Winnipeg, Manitoba, Canada, 2 Departments of Pediatrics, Physiology, University of Manitoba, Winnipeg, Manitoba, Canada, 3 Biology of Breathing Group- Manitoba Institute of Child Health, Winnipeg, Manitoba, Canada, 4 Department of Internal Medicine (Cardiology), Cardiovascular Research Center, Yale University School of Medicine, New Haven, Connecticut, United States of America Abstract G protein-coupled receptors (GPCRs) show some level of basal activity even in the absence of an agonist, a phenomenon referred to as constitutive activity. Such constitutive activity in GPCRs is known to have important pathophysiological roles in human disease. The thromboxane A2 receptor (TP) is a GPCR that promotes thrombosis in response to binding of the prostanoid, thromboxane A2. TP dysfunction is widely implicated in pathophysiological conditions such as bleeding disorders, hypertension and cardiovascular disease. Recently, we reported the characterization of a few constitutively active mutants (CAMs) in TP, including a genetic variant A160T. Using these CAMs as reporters, we now test the inverse agonist properties of known antagonists of TP, SQ 29,548, Ramatroban, L-670596 and Diclofenac, in HEK293T cells. Interestingly, SQ 29,548 reduced the basal activity of both, WT-TP and the CAMs while Ramatroban was able to reduce the basal activity of only the CAMs. Diclofenac and L-670596 showed no statistically significant reduction in basal activity of WT-TP or CAMs. To investigate the role of these compounds on human platelet function, we tested their effects on human megakaryocyte based system for platelet activation. -

G-Protein-Coupled Receptor Signaling and Polarized Actin Dynamics Drive
RESEARCH ARTICLE elifesciences.org G-protein-coupled receptor signaling and polarized actin dynamics drive cell-in-cell invasion Vladimir Purvanov, Manuel Holst, Jameel Khan, Christian Baarlink, Robert Grosse* Institute of Pharmacology, University of Marburg, Marburg, Germany Abstract Homotypic or entotic cell-in-cell invasion is an integrin-independent process observed in carcinoma cells exposed during conditions of low adhesion such as in exudates of malignant disease. Although active cell-in-cell invasion depends on RhoA and actin, the precise mechanism as well as the underlying actin structures and assembly factors driving the process are unknown. Furthermore, whether specific cell surface receptors trigger entotic invasion in a signal-dependent fashion has not been investigated. In this study, we identify the G-protein-coupled LPA receptor 2 (LPAR2) as a signal transducer specifically required for the actively invading cell during entosis. We find that 12/13G and PDZ-RhoGEF are required for entotic invasion, which is driven by blebbing and a uropod-like actin structure at the rear of the invading cell. Finally, we provide evidence for an involvement of the RhoA-regulated formin Dia1 for entosis downstream of LPAR2. Thus, we delineate a signaling process that regulates actin dynamics during cell-in-cell invasion. DOI: 10.7554/eLife.02786.001 Introduction Entosis has been described as a specialized form of homotypic cell-in-cell invasion in which one cell actively crawls into another (Overholtzer et al., 2007). Frequently, this occurs between tumor cells such as breast, cervical, or colon carcinoma cells and can be triggered by matrix detachment (Overholtzer et al., 2007), suggesting that loss of integrin-mediated adhesion may promote cell-in-cell invasion. -

G-Protein-Coupled Receptor Heteromer Dynamics
Commentary 4215 G-protein-coupled receptor heteromer dynamics Jean-Pierre Vilardaga1,2,*, Luigi F. Agnati3, Kjell Fuxe4 and Francisco Ciruela5 1Laboratory for GPCR Biology, Department of Pharmacology and Chemical Biology, University of Pittsburgh, School of Medicine, Pittsburgh, PA 15261, USA 2Endocrine Unit, Department of Medicine, Massachusetts General Hospital and Harvard Medical School, Boston, MA 02114, USA 3IRCCS, San Camillo, Lido Venezia, Italy 4Department of Neuroscience, Karolinska Institute, Stockholm SE-17177, Sweden 5Pharmacology Unit, Department of Pathology and Experimental Therapy, Faculty of Medicine, University of Barcelona, 08907 Barcelona, Spain *Author for correspondence ([email protected]) Journal of Cell Science 123, 000-000 © 2010. Published by The Company of Biologists Ltd doi:10.1242/jcs.063354 Summary G-protein-coupled receptors (GPCRs) represent the largest family of cell surface receptors, and have evolved to detect and transmit a large palette of extracellular chemical and sensory signals into cells. Activated receptors catalyze the activation of heterotrimeric G proteins, which modulate the propagation of second messenger molecules and the activity of ion channels. Classically thought to signal as monomers, different GPCRs often pair up with each other as homo- and heterodimers, which have been shown to modulate signaling to G proteins. Here, we discuss recent advances in GPCR heteromer systems involving the kinetics of the early steps in GPCR signal transduction, the dynamic property of receptor–receptor interactions, and how the formation of receptor heteromers modulate the kinetics of G-protein signaling. Key words: G-protein-coupled receptors, Heterodimers, Signaling Introduction the signaling and trafficking mechanisms of GPCRs is thus central G-protein-coupled receptors (GPCRs) constitute the main family for the development of new and safer therapies for many of cell surface receptors for a large variety of chemical stimuli physiological and psychological disorders. -
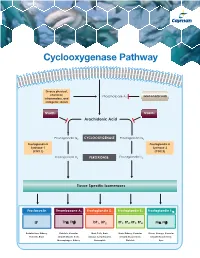
Cyclooxygenase Pathway
Cyclooxygenase Pathway Diverse physical, chemical, Phospholipase A Glucocorticoids inflammatory, and 2 mitogenic stimuli NSAIDs NSAIDs Arachidonic Acid Prostaglandin G2 CYCLOOXYGENASE Prostaglandin G2 Prostaglandin H Prostaglandin H Synthase-1 Synthase-2 (COX 1) (COX 2) Prostaglandin H2 PEROXIDASE Prostaglandin H2 Tissue Specific Isomerases Prostacyclin Thromboxane A2 Prostaglandin D2 Prostaglandin E2 Prostaglandin F2α IP TPα, TPβ DP1, DP2 EP1, EP2, EP3, EP4 FPα, FPβ Endothelium, Kidney, Platelets, Vascular Mast Cells, Brain, Brain, Kidney, Vascular Uterus, Airways, Vascular Platelets, Brain Smooth Muscle Cells, Airways, Lymphocytes, Smooth Muscle Cells, Smooth Muscle Cells, Macrophages, Kidney Eosinophils Platelets Eyes Prostacyclin Item No. Product Features Prostacyclin (Prostaglandin I2; PGI2) is formed from arachidonic acid primarily in the vascular endothelium and renal cortex by sequential 515211 6-keto • Sample Types: Culture Medium | Plasma Prostaglandin • Measure 6-keto PGF levels down to 6 pg/ml activities of COX and prostacyclin synthase. PGI2 is non-enzymatically 1α F ELISA Kit • Incubation : 18 hours | Development: 90-120 minutes | hydrated to 6-keto PGF1α (t½ = 2-3 minutes), and then quickly converted 1α Read: Colorimetric at 405-420 nm to the major metabolite, 2,3-dinor-6-keto PGF1α (t½= 30 minutes). Prostacyclin was once thought to be a circulating hormone that regulated • Assay 24 samples in triplicate or 36 samples in duplicate platelet-vasculature interactions, but the rate of secretion into circulation • NOTE: A portion of urinary 6-keto PGF1α is of renal origin coupled with the short half-life indicate that prostacyclin functions • NOTE : It has been found that normal plasma levels of 6-keto PGF may be low locally. -

Vasoactive Responses of U46619, PGF2 , Latanoprost, and Travoprost
Vasoactive Responses of U46619, PGF2␣, Latanoprost, and Travoprost in Isolated Porcine Ciliary Arteries Ineta Vysniauskiene, Reto Allemann, Josef Flammer, and Ivan O. Haefliger PURPOSE. To compare the vasoactive properties of the prosta- these responses can be modulated by SQ 29548 (TP-receptor noids U46619 (thromboxane A2 analogue), prostaglandin F2␣ antagonist) or AL-8810 (FP-receptor antagonist). (PGF2␣), latanoprost free acid, and travoprost free acid in isolated porcine ciliary arteries. METHODS. In a myograph system (isometric force measure- MATERIAL AND METHODS ment), quiescent vessels were exposed (cumulatively) to U46619, PGF , latanoprost, or travoprost (0.1 nM–0.1 mM). 2␣ Vessels Preparation Experiments were also conducted in the presence of SQ 29548 (TP-receptor antagonist; 3–10 M) or AL-8810 (FP-receptor Porcine eyes were obtained from an abattoir immediately after death. antagonist; 3–30 M). Contractions were expressed as the In cold modified Krebs-Ringer solution (NaCl 118 mM, KCl 4.7 mM, percentage of 100 mM potassium chloride-induced contrac- CaCl2 2.5 mM, K2PO4 1.2 mM, MgSO4 1.2 mM, NaHCO3 25 mM, tions. glucose 11.1 mM, and EDTA 0.026 mM), ciliary arteries were dissected 5 RESULTS. In quiescent vessels, contractions (concentration–re- and cut into 2-mm segments. In an organ chamber, two 45- m Ϯ tungsten wires were passed through the vessel’s lumen and attached to sponse curves) induced by (0.1 mM) PGF2␣ (87.9% 3.5%), U46619 (66.7% Ϯ 4.1%), and latanoprost (62.9% Ϯ 3.6%) were a force transducer for isometric force measurements (Myo-Interface; JP more pronounced (P Յ 0.001) than those induced by tra- Trading, Aarhus, Denmark). -

G Protein-Coupled Receptors: What a Difference a ‘Partner’ Makes
Int. J. Mol. Sci. 2014, 15, 1112-1142; doi:10.3390/ijms15011112 OPEN ACCESS International Journal of Molecular Sciences ISSN 1422-0067 www.mdpi.com/journal/ijms Review G Protein-Coupled Receptors: What a Difference a ‘Partner’ Makes Benoît T. Roux 1 and Graeme S. Cottrell 2,* 1 Department of Pharmacy and Pharmacology, University of Bath, Bath BA2 7AY, UK; E-Mail: [email protected] 2 Reading School of Pharmacy, University of Reading, Reading RG6 6UB, UK * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +44-118-378-7027; Fax: +44-118-378-4703. Received: 4 December 2013; in revised form: 20 December 2013 / Accepted: 8 January 2014 / Published: 16 January 2014 Abstract: G protein-coupled receptors (GPCRs) are important cell signaling mediators, involved in essential physiological processes. GPCRs respond to a wide variety of ligands from light to large macromolecules, including hormones and small peptides. Unfortunately, mutations and dysregulation of GPCRs that induce a loss of function or alter expression can lead to disorders that are sometimes lethal. Therefore, the expression, trafficking, signaling and desensitization of GPCRs must be tightly regulated by different cellular systems to prevent disease. Although there is substantial knowledge regarding the mechanisms that regulate the desensitization and down-regulation of GPCRs, less is known about the mechanisms that regulate the trafficking and cell-surface expression of newly synthesized GPCRs. More recently, there is accumulating evidence that suggests certain GPCRs are able to interact with specific proteins that can completely change their fate and function. These interactions add on another level of regulation and flexibility between different tissue/cell-types. -

Effect of Prostanoids on Human Platelet Function: an Overview
International Journal of Molecular Sciences Review Effect of Prostanoids on Human Platelet Function: An Overview Steffen Braune, Jan-Heiner Küpper and Friedrich Jung * Institute of Biotechnology, Molecular Cell Biology, Brandenburg University of Technology, 01968 Senftenberg, Germany; steff[email protected] (S.B.); [email protected] (J.-H.K.) * Correspondence: [email protected] Received: 23 October 2020; Accepted: 23 November 2020; Published: 27 November 2020 Abstract: Prostanoids are bioactive lipid mediators and take part in many physiological and pathophysiological processes in practically every organ, tissue and cell, including the vascular, renal, gastrointestinal and reproductive systems. In this review, we focus on their influence on platelets, which are key elements in thrombosis and hemostasis. The function of platelets is influenced by mediators in the blood and the vascular wall. Activated platelets aggregate and release bioactive substances, thereby activating further neighbored platelets, which finally can lead to the formation of thrombi. Prostanoids regulate the function of blood platelets by both activating or inhibiting and so are involved in hemostasis. Each prostanoid has a unique activity profile and, thus, a specific profile of action. This article reviews the effects of the following prostanoids: prostaglandin-D2 (PGD2), prostaglandin-E1, -E2 and E3 (PGE1, PGE2, PGE3), prostaglandin F2α (PGF2α), prostacyclin (PGI2) and thromboxane-A2 (TXA2) on platelet activation and aggregation via their respective receptors. Keywords: prostacyclin; thromboxane; prostaglandin; platelets 1. Introduction Hemostasis is a complex process that requires the interplay of multiple physiological pathways. Cellular and molecular mechanisms interact to stop bleedings of injured blood vessels or to seal denuded sub-endothelium with localized clot formation (Figure1).