Habitat Selection and Calling Activity of the Lewin's Rail (Lewinia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Classification of the Rallidae
A CLASSIFICATION OF THE RALLIDAE STARRY L. OLSON HE family Rallidae, containing over 150 living or recently extinct species T and having one of the widest distributions of any family of terrestrial vertebrates, has, in proportion to its size and interest, received less study than perhaps any other major group of birds. The only two attempts at a classifi- cation of all of the recent rallid genera are those of Sharpe (1894) and Peters (1934). Although each of these lists has some merit, neither is satisfactory in reflecting relationships between the genera and both often separate closely related groups. In the past, no attempt has been made to identify the more primitive members of the Rallidae or to illuminate evolutionary trends in the family. Lists almost invariably begin with the genus Rdus which is actually one of the most specialized genera of the family and does not represent an ancestral or primitive stock. One of the difficulties of rallid taxonomy arises from the relative homo- geneity of the family, rails for the most part being rather generalized birds with few groups having morphological modifications that clearly define them. As a consequence, particularly well-marked genera have been elevated to subfamily rank on the basis of characters that in more diverse families would not be considered as significant. Another weakness of former classifications of the family arose from what Mayr (194933) referred to as the “instability of the morphology of rails.” This “instability of morphology,” while seeming to belie what I have just said about homogeneity, refers only to the characteristics associated with flightlessness-a condition that appears with great regularity in island rails and which has evolved many times. -

A Synopsis of the Pre-Human Avifauna of the Mascarene Islands
– 195 – Paleornithological Research 2013 Proceed. 8th Inter nat. Meeting Society of Avian Paleontology and Evolution Ursula B. Göhlich & Andreas Kroh (Eds) A synopsis of the pre-human avifauna of the Mascarene Islands JULIAN P. HUME Bird Group, Department of Life Sciences, The Natural History Museum, Tring, UK Abstract — The isolated Mascarene Islands of Mauritius, Réunion and Rodrigues are situated in the south- western Indian Ocean. All are volcanic in origin and have never been connected to each other or any other land mass. Despite their comparatively close proximity to each other, each island differs topographically and the islands have generally distinct avifaunas. The Mascarenes remained pristine until recently, resulting in some documentation of their ecology being made before they rapidly suffered severe degradation by humans. The first major fossil discoveries were made in 1865 on Mauritius and on Rodrigues and in the late 20th century on Réunion. However, for both Mauritius and Rodrigues, the documented fossil record initially was biased toward larger, non-passerine bird species, especially the dodo Raphus cucullatus and solitaire Pezophaps solitaria. This paper provides a synopsis of the fossil Mascarene avifauna, which demonstrates that it was more diverse than previously realised. Therefore, as the islands have suffered severe anthropogenic changes and the fossil record is far from complete, any conclusions based on present avian biogeography must be viewed with caution. Key words: Mauritius, Réunion, Rodrigues, ecological history, biogeography, extinction Introduction ily described or illustrated in ships’ logs and journals, which became the source material for The Mascarene Islands of Mauritius, Réunion popular articles and books and, along with col- and Rodrigues are situated in the south-western lected specimens, enabled monographs such as Indian Ocean (Fig. -

A Trait Dataset for Taiwan's Breeding Birds
Biodiversity Data Journal 8: e49735 doi: 10.3897/BDJ.8.e49735 Data Paper A trait dataset for Taiwan's breeding birds Pei-Yu Tsai‡, Chie-Jen Ko §,|, Chia Hsieh¶#, Yi-Ting Su , Ya-Jung Lu‡, Ruey-Shing Lin§, Mao-Ning Tuanmu¤ ‡ Biodiversity Research Center, Academia Sinica, Taipei, Taiwan § Endemic Species Research Institute, Jiji, Nantou, Taiwan | Institute of Ecology and Evolutionary Biology, National Taiwan University, Taipei, Taiwan ¶ BioSciences Department, Rice University, Houston, United States of America # Department of Life Sciences, National Cheng Kung University, Tainan, Taiwan ¤ Thematic Center for Systematics and Biodiversity Informatics, Biodiversity Research Center, Academia Sinica, Taipei, Taiwan Corresponding author: Mao-Ning Tuanmu ([email protected]) Academic editor: Cynthia Parr Received: 30 Dec 2019 | Accepted: 08 May 2020 | Published: 19 May 2020 Citation: Tsai P-Y, Ko C-J, Hsieh C, Su Y-T, Lu Y-J, Lin R-S, Tuanmu M-N (2020) A trait dataset for Taiwan's breeding birds. Biodiversity Data Journal 8: e49735. https://doi.org/10.3897/BDJ.8.e49735 Abstract Background Species traits affect how a species interacts with the environment and other species and thus determine the role of the species in an ecosystem. They affect not only population dynamics of a species across space and over time, but also community structure and function through their key role in the community assembly processes. Information on species traits is also highly relevant for conservation planning as they determine the adaptive ability of a species in the face of environmental changes. However, information on species traits is usually scarce and sparsely distributed amongst diverse types of literature and sources. -

ON the NATIVE FAUNA of ALDABRA ATOLL, SEYCHELLES Ferai Cats
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by I-Revues THE IMPACT OF THE FERAL CAT (FELIS CATUS) ON THE NATIVE FAUNA OF ALDABRA ATOLL, SEYCHELLES Wendy SEABROOK* Ferai cats (Fe lis catus) have colonized and caused considerable damage to island ecosystems throughout the world (Merton, 1978 ; Croxall et al., 1984 ; Veitch, 1985). In this paper the results of an examination (1985-86) of the distribution and ecology of the ferai cat on Aldabra Atoll in the Indian Ocean (9.5" S., 46.5" E.) are summarized, and the impact of cats on native fauna is assessed. Prior to this study very little was known about the cat population. Only sightings and signs of them had been reported on the four main islands of the Atoll (Stoddart, 1971 ; Racey Nicoll, 1984). Considering Aldabra's status as a World Heritage Site and its rich endemie fauna (eg. Giant tortoise (Geochelone& gigantea) and White-throated rail ( Dryo limnas cuvieri aldabranus)), a preliminary ecological study on the cats was necessary to design an eradication or control programme. Any proposai to eliminate an alien predator from an island should be justified on biological grounds, and the likely ecological consequences must be considered (Merton, 1978), especially if, as on Aldabra, other alien predators are also present. Because of the considerable financial investment and persan-power required, eradication and control programmes must also be evaluated with reference to other mana gement objectives. Aldabra is one of the finest examples in the world of a raised coral atoll ecosystem (Stoddart Savy, 1983). -

Archiv Für Naturgeschichte
© Biodiversity Heritage Library, http://www.biodiversitylibrary.org/; www.zobodat.at Bericht über die Leistungen in der Naturgeschichte der Vögel während des Jahres 1896. Von C. B. Hellmayr. I. Allgemeines, Sammlungen und Museen, Personalien, Reisen, Taxidermie. Sir Joseph Banks. Journal of the Rt. Hon. Sir Joseph Banks, during Capt. Cook's First Voyage in H. M. S. „Endeavour" in 1768 —71 to Tierra del Fiiego, Otaheite, New Zealand, Australia, the Dutch East Indies etc. Edited by Sir Joseph D. Hooker. London. 1896. 8'\ LIX und 466 pg. J. Banks begleitete Capt. Cook auf dessen erster Weltumseglung als Naturforscher. Den Inhalt des Buches bildet das Reisetagebuch, das viele eingestreute Beobachtungen über die angetroffenen Vögel enthält. Der Herausgeber fügt eine biographische Skizze des Ver- fassers bei. Porträts von J. Banks und seinem Gehilfen Solander sind dem Buche beigegeben. H. E. Barnes, Nekrolog; Ibis (7) IL p. 162. R. Baron et P. Dechambre. De l'espece et des races chez les animaux superieurs et specialement de leurs rapports avec le polvmorphisme sexuel: Bull. Scientif. France, Belg. XXVII pt. II (1896) p. 338—356. Bericht über die Reise der Herren Lauterbach, Kersting a. Tappenbeck: Orn. Monber. IV p. 151. A. P. Bogdauow. Todesanzeige, von Paul Leverkühn; Orn. Monatsschr. Schutz Vogelw. XXI (1896) p. 229-230. E. T. Booth. Catalogue of the Cases of Birds in the Dyke Road Museum, Brighton, 2. edit. (by A. F. Griffith). With 16 füll page illustr. Brighton 1896. 8''. 217*^8. Braunschweiger naturhistorisches Museum. Orn ^Ib. IV. p. 66—67. Robert Brown, Nekrolog; Ibis (7) II p. 163. Aixh. f. -
Eton Range Realignment Project ATTACHMENT 2 to EPBC Ref: 2015/7552 Preliminary Documentation Residual Impact Assessment and Offset Proposal - 37
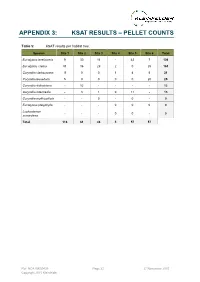
APPENDIX 3: KSAT RESULTS – PELLET COUNTS Table 5: KSAT results per habitat tree. Species Site 1 Site 2 Site 3 Site 4 Site 5 Site 6 Total Eucalyptus tereticornis 9 30 16 - 42 7 104 Eucalyptus crebra 91 16 29 2 0 25 163 Corymbia clarksoniana 11 0 0 1 4 5 21 Corymbia tessellaris 5 0 0 0 0 20 25 Corymbia dallachiana - 12 - - - - 12 Corymbia intermedia - 3 1 0 11 - 15 Corymbia erythrophloia - - 0 - 0 - 0 Eucalyptus platyphylla - - - 0 0 0 0 Lophostemon - - - 0 0 - 0 suaveolens Total 116 61 46 3 57 57 Ref: NCA15R30439 Page 22 27 November 2015 Copyright 2015 Kleinfelder APPENDIX 4: SITE PHOTOS The following images were taken from the centre of each BioCondition quadrat and represent a north east south west aspect, top left to bottom right. Ref: NCA15R30439 Page 23 27 November 2015 Copyright 2015 Kleinfelder Plate 3: BioCondition quadrat 1 (RE11.3.4/11.12.3) Ref: NCA15R30439 Page 24 27 November 2015 Copyright 2015 Kleinfelder Plate 4: BioCondition quadrat 2 (RE11.3.4/11.12.3) Ref: NCA15R30439 Page 25 27 November 2015 Copyright 2015 Kleinfelder Plate 5: BioCondition quadrat 3 (RE11.12.3) Ref: NCA15R30439 Page 26 27 November 2015 Copyright 2015 Kleinfelder Plate 6: BioCondition quadrat 4 (RE11.3.9) Ref: NCA15R30439 Page 27 27 November 2015 Copyright 2015 Kleinfelder Plate 7: BioCondition quadrat 5 (RE11.3.25) Ref: NCA15R30439 Page 28 27 November 2015 Copyright 2015 Kleinfelder Plate 8: BioCondition quadrat 6 (RE11.12.3/11.3.4/11.3.9) Ref: NCA15R30439 Page 29 27 November 2015 Copyright 2015 Kleinfelder Appendix E: Desktop Assessment for Potential -

Index to Volume 85, 1973
INDEX TO VOLUME 85, 1973 BY EMMA J. MESSERLY, JOHN F. MESSERLY AND MARY M. MCCLELLAN This index includes in addition to the names of genera, species, and authors, references to the following topics: abnormalities, aggression, anatomy, banding, behavior, brood size, care of young, clutch size, communication, conservation, courtship, density, display, distri- bution, ecology, eggs, food and food habits, growth and development, habitat, hatching, incubation, locomotion, measurements, metabolism, migration, molts and plumages, mor- phology, mortality, nest building, nest sites, nesting, nesting failure, nesting success, nests, parasitism, pesticides, physiology, populations, predation, roosting, taxonomy, territory, voice and vocalizations, weather, and weights. Also included are references of biological significance to reptiles and mammals. Forms described as new in this volume are given in boldface. Abnormality, 329, 34&342, 345, 346 akool, 403, 404 Acanthis hornemanni, 484 isabellina, 403, 408 Actitis hypoleucos, 143 olivaceus, 403, 408 macnlaria, 143, 473 olivieri, 404 Acrocephalus aequinoctialis, 95 phoenicurus, 403, 404, 409 familiaris kingi, 90 Amazon, 494 Aechmophoros occidentalis, 449 Orange-winged, 494 Aenigmatolimnas, 410, 411 Red-crowned, 494 marginalis, 404 White-fronted, 494 Afropavo, 390 Amazona, 509 Agapornis personata, 503 albifrons, 494 Agelaius phoeniceus, 86, 209, 342, 450, 483 amazonica, 494 Aggression, 22, 23-24, 63-64, 71-74, 78, autumnalis, 509 166, 289, 332-334, 338-339, 342-343, ochrocephala, 502, 508, 509 439, 440, 4499451, 40-481 Aimophila aestivalis, 50 viridigenalis, 494, 509 a. aestivalis, 50 Amblycercus holosericeus, 240, 241 a. bachmanii, 50 Ammodramus savannarum, 50 ruficeps, 346 s. pratensis, 50 Aix sponsa, 308, 467, 469 Ammospiza caudacuta, 50 Akahige, Ryukyu, 16 c. nelsoni, 50 Albatross, Black-footed, 91, 92 maritima, 348 Laysan, 91, 92, 94, 332 Amphispiza belli, 348 Wandering, 331 Anarhynchus frontalis, 61, 68, 70 Alcock, John, see Bird, Joan, and ___ Anas, 297, 298, 302, 303, 304 Alectoris chukar. -

Supplementary Methods S1
1 Validation methods for trophic niche models 2 3 To assign links between nodes (species), we used trophic niche-space models (e.g., [1]). 4 Each of these models has two quantile regressions that define the prey-size range a 5 predator of a given size is predicted to consume. Species whose body mass is within the 6 range of a predator’s prey size, as identified by the trophic niche-space model, are predicted 7 to be prey, while those outside the range are predicted not to be eaten. 8 9 The broad taxonomy of a predator helps to predict predation interactions [2]. To optimize 10 our trophic niche-space model, we therefore tested whether including taxonomic class of 11 predators improved the fit of quantile regressions. Using trophic (to identify which species 12 were predators), body mass, and taxonomic data, we fitted and compared five quantile 13 regression models (including a null model) to the GloBI data. In each model, we log10- 14 transformed the dependent variable prey body mass, and included for the independent 15 variables different combinations of log10-transformed predator body mass, predator class, 16 and the interaction between these variables (Supplementary Table S4). We log10- 17 transformed both predator and prey body mass to linearize the relationship between these 18 variables. We fit the five quantile regressions to the upper and lower 5% of prey body mass, 19 and compared model fits using the Bayesian information criterion (BIC). The predator body 20 mass*predator class model fit the 95th quantile data best, whereas the predator body mass 21 + predator class model fit the 5th quantile data marginally better than the aforementioned 22 interaction model (Supplementary Figure S2, Supplementary Table S4). -

Standards for Aquatic/Semi-Aquatic Bird Sanctuaries
Global Federation of Animal Sanctuaries Standards For Aquatic/Semi-Aquatic Bird Sanctuaries Version: April 2019 ©2012 Global Federation of Animal Sanctuaries Global Federation of Animal Sanctuaries – Standards for Aquatic/Semi-Aquatic Bird Sanctuaries Table of Contents INTRODUCTION ............................................................................................................ 1 GFAS PRINCIPLES ............................................................................................................................................................. 1 ANIMALS COVERED BY THESE STANDARDS ................................................................................................. 1 AQUATIC/SEMI-AQUATIC BIRD STANDARDS ............................................................................................. 3 AQUATIC/SEMI-AQUATIC BIRD HOUSING ....................................................... 3 H-1. Types of Space and Size .................................................................................................................................................... 3 H-2. Containment ................................................................................................................................................................................ 5 H-3. Ground and Plantings ........................................................................................................................................................... 6 H-4. Gates and Doors ...................................................................................................................................................................... -

Download Article As 589.6 KB PDF File
6 AvailableNew on-lineZealand at: Journal http://www.newzealandecology.org/nzje/ of Ecology, Vol. 34, No. 1, 2010 special issue: Feathers to Fur The ecological transformation of Aotearoa/New Zealand The origin and history of New Zealand’s terrestrial vertebrates Alan J.D. Tennyson Museum of New Zealand Te Papa Tongarewa, PO Box 467, Wellington, New Zealand (Email: [email protected]) Published on-line: 4 November 2009 Abstract: Since the 1980s, morphological and molecular research has resulted in significant advances in understanding the relationships and origins of the recent terrestrial vertebrate fauna in the New Zealand biogeographic region. This research has led to many taxonomic changes, with a significant increase in the number of bird and reptile species recognised. It has also resulted in the recognition of several more Holocene (<10 000 years ago) bird species extinctions. The conclusion that Holocene extinctions were primarily caused by human- hunting and predation by other introduced mammals (particularly rats and cats) has been supported by new data. Despite many local eradications of introduced pests, the number of introduced species has increased, with the establishment of five more foreign birds and (on Norfolk Island) the house gecko (Hemidactylus frenatus). Many new, significant New Zealand vertebrate fossils have been reported, including more dinosaurs from the Cretaceous, and the first Tertiary records of frogs, rhynchocephalids, lizards, crocodylians, bats and a terrestrial “Mesozoic ghost” mammal from the Early Miocene near St Bathans. For birds, the earliest known penguins in the world have been discovered, and there are intriguing Late Cretaceous – Early Paleocene remains still awaiting detailed description. -

Supporting References for Nelson & Ellis
Supplemental Data for Nelson & Ellis (2018) The citations below were used to create Figures 1 & 2 in Nelson, G., & Ellis, S. (2018). The History and Impact of Digitization and Digital Data Mobilization on Biodiversity Research. Publication title by year, author (at least one ADBC funded author or not), and data portal used. This list includes papers that cite the ADBC program, iDigBio, TCNs/PENs, or any of the data portals that received ADBC funds at some point. Publications were coded as "referencing" ADBC if the authors did not use portal data or resources; it includes publications where data was deposited or archived in the portal as well as those that mention ADBC initiatives. Scroll to the bottom of the document for a key regarding authors (e.g., TCNs) and portals. Citation Year Author Portal used Portal or ADBC Program was referenced, but data from the portal not used Acevedo-Charry, O. A., & Coral-Jaramillo, B. (2017). Annotations on the 2017 Other Vertnet; distribution of Doliornis remseni (Cotingidae ) and Buthraupis macaulaylibrary wetmorei (Thraupidae ). Colombian Ornithology, 16, eNB04-1 http://asociacioncolombianadeornitologia.org/wp- content/uploads/2017/11/1412.pdf [Accessed 4 Apr. 2018] Adams, A. J., Pessier, A. P., & Briggs, C. J. (2017). Rapid extirpation of a 2017 Other VertNet North American frog coincides with an increase in fungal pathogen prevalence: Historical analysis and implications for reintroduction. Ecology and Evolution, 7, (23), 10216-10232. Adams, R. P. (2017). Multiple evidences of past evolution are hidden in 2017 Other SEINet nrDNA of Juniperus arizonica and J. coahuilensis populations in the trans-Pecos, Texas region. -

Diversity and Endemism in Tidal-Marsh Vertebrates
Studies in Avian Biology No. 32:32-53 DIVERSITY AND ENDEMISM IN TIDAL-MARSH VERTEBRATES RUSSELL GREENBERG AND JESúS E. MALDONADO Abstract. Tidal marshes are distributed patcliily, predominantly along the mid- to high-latitude coasts of the major continents. The greatest extensions of non-arctic tidal marshes are found along the Atlantic and Gulf coasts of North America, but local concentrations can be found in Great Britain, northern Europe, northern Japan, northern China, and northern Korea, Argentina-Uruguay-Brazil, Australia, and New Zealand. We tallied the number of terrestrial vertebrate species that regularly occupy tidal marshes in each of these regions, as well as species or subspecies that are largely restricted to tidal marshes. In each of the major coastal areas we found 8-21 species of breeding birds and 13-25 species of terrestrial mammals. The diversity of tidal-marsh birds and mammals is highly inter-correlated, as is the diversity of species restricted to saltmarshes. These values are, in turn, correlated with tidal-marsh area along a coastline. We estimate approximately seven species of turtles occur in brackish or saltmarshes worldwide, but only one species is endemic and it is found in eastern North America. A large number of frogs and snakes occur opportunistically in tidal marshes, primarily in southeastern United States, particularly Florida. Three endemic snake taxa are restricted to tidal marshes of eastern North America as well. Overall, only in North America were we able to find documentation for multiple taxa of terrestrial vertebrates associated with tidal marshes. These include one species of mammal and two species of birds, one species of snake, and one species of turtle.