Partridge and Embryonated Partridge Egg As New Preclinical Models For
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Printable PDF Format
Field Guides Tour Report Taiwan 2020 Feb 1, 2020 to Feb 12, 2020 Phil Gregory & local guide Arco Huang For our tour description, itinerary, past triplists, dates, fees, and more, please VISIT OUR TOUR PAGE. This gorgeous male Swinhoe's Pheasant was one of the birds of the trip! We found a pair of these lovely endemic pheasants at Dasyueshan. Photo by guide Phil Gregory. This was a first run for the newly reactivated Taiwan tour (which we last ran in 2006), with a new local organizer who proved very good and enthusiastic, and knew the best local sites to visit. The weather was remarkably kind to us and we had no significant daytime rain, somewhat to my surprise, whilst temperatures were pretty reasonable even in the mountains- though it was cold at night at Dasyueshan where the unheated hotel was a bit of a shock, but in a great birding spot, so overall it was bearable. Fog on the heights of Hohuanshan was a shame but at least the mid and lower levels stayed clear. Otherwise the lowland sites were all good despite it being very windy at Hengchun in the far south. Arco and I decided to use a varied assortment of local eating places with primarily local menus, and much to my amazement I found myself enjoying noodle dishes. The food was a highlight in fact, as it was varied, often delicious and best of all served quickly whilst being both hot and fresh. A nice adjunct to the trip, and avoided losing lots of time with elaborate meals. -

Macrohabitat Characteristics and Distribution Hotspots of Endemic Bird Species in Taiwan
Taiwania, 55(3): 216-227, 2010 Macrohabitat Characteristics and Distribution Hotspots of Endemic Bird Species in Taiwan Chia-Ying Ko(1*), Ruey-Shing Lin(2) and Pei-Fen Lee(1) 1. Institute of Ecology and Evolutionary Biology, National Taiwan University, 1, Roosevelt Rd., Sec. 4, Taipei 106, Taiwan; equal contribution. 2. Endemic Species Research Institute, 1, Ming-Shen E. Road, Jiji, Nantou 552, Taiwan. * Corresponding author. Tel: 866-2-3366246; Fax: 886-2-23623501; Email: [email protected] (Manuscript received 24 February 2010; accepted 23 April 2010) ABSTRACT: Understanding species distributions is essential for developing biodiversity conservation strategies. We combined two bird inventories conducted from 1993 to 2004 and identified specific features of 17 endemic bird species in Taiwan. We used eight environmental variables, including elevation, annual total precipitation, annual mean temperature, warmth index, percentage of forest cover, mean Normalized Difference Vegetation Index (NDVI), percentage of building area, and road density, to define macrohabitat characteristics of each species. All the data were in a 1 × 1 km gird system. The 17 species were classified as common (being present in more than 200 grids), uncommon (100–200 grids) or rare (less than 100 grids). The Mikado Pheasant (Syrmaticus mikado), as a rare species, had the lowest occurrence records, while the Taiwan Barbet (Megalaima nuchalis), as a common species, had the highest. Each species had a specific distribution range and habitat preference. These 17 species occupied heterogeneous elevation and climatic conditions. In general, they favored habitats with high vegetation cover, at almost full forest cover and median to high NDVI. Canonical correspondence analysis (CCA) indicated that elevation had the highest correlations with species distributions, with axis 1 accounting for 57.5% of the variation and axis 2 for 9.8%. -

A Global Study of the Distribution and Richness of Alien Bird Species
A global study of the distribution and richness of alien bird species Ellie E. Dyer A dissertation submitted for the degree of Doctor of Philosophy University College London Centre for Biodiversity and Environment Research (CBER) within the Department of Genetics, Evolution and Environment (GEE) University College London November 4, 2015 Declaration I, Ellie Eveness Dyer, confirm that the work presented in this thesis is my own. Where information has been derived from other sources, I confirm that this has been indicated in the thesis. Ellie Dyer, 4th November 2015 2 Abstract Alien species are a major component of human-induced environmental change, yet spatial and temporal variation in the drivers of their introduction, and their subsequent distribution and richness, are poorly understood. Here, I present a global analysis of the drivers of this variation for a major animal group, birds (Class Aves), using the newly-created Global Avian Invasions Atlas (GAVIA) database. GAVIA includes information on introduction successes and failures, enabling me to examine the effect of colonisation pressure (the number of species introduced) on alien bird distributions. A description of the GAVIA database is given in Chapter 2, with details on its scope and sources, data collation and validation, and the production of alien range maps. Chapter 3 focuses on the early stages of the invasion pathway, and shows that historical introductions tend to originate in Europe, were driven by the global movements of British colonialism, and involved species deemed useful. Modern introductions, in contrast, tend to originate in Southeast Asia and Africa, are driven by factors associated with wealth, and involve species found in the pet trade. -

Is the Clouded Leopard Neofelis Nebulosa Extinct in Taiwan, and Could It Be Reintroduced? an Assessment of Prey and Habitat
Is the clouded leopard Neofelis nebulosa extinct in Taiwan, and could it be reintroduced? An assessment of prey and habitat P O -JEN C HIANG,KURTIS J AI-CHYI P EI,MICHAEL R. VAUGHAN C HING-FENG L I ,MEI-TING C HEN,JIAN-NAN L IU,CHUNG-YI L IN L IANG-KONG L IN and Y U -CHING L AI Abstract During – we conducted a nationwide become over-abundant. Thus, it is important to address camera-trapping survey and assessed the availability of the cascading effect of the disappearance of top-down pred- prey and habitat for the clouded leopard Neofelis nebulosa ator control. Our assessment indicated that, with proper in Taiwan. We surveyed , camera-trap sites over regulation of hunting, habitat restoration and corridor im- , camera-trap days, from the seashore to an altitude provement, it may be possible to reintroduce the clouded of , m and covering various types of vegetation. No leopard. clouded leopards were photographed during , cam- Keywords Camera-trapping, extinct, habitat assessment, era-trap days, including at sites in other studies, con- Neofelis nebulosa, prey distribution, reintroduction, Taiwan firming the presumed extinction of clouded leopards in Taiwan. Assessment of the prey base revealed altitudinal distribution patterns of prey species and prey biomass. Areas at lower altitudes and with less human encroachment and hunting supported a higher prey biomass and more of Introduction the typical prey species of clouded leopards. Habitat analysis he clouded leopard Neofelis nebulosa is categorized as revealed , km of suitable habitat but this was reduced to Vulnerable on the IUCN Red List and is listed on T , km when adjacent areas of human encroachment Appendix I of CITES (). -
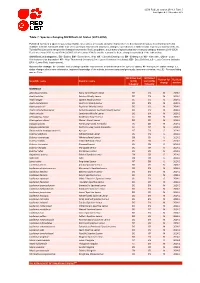
Table 7: Species Changing IUCN Red List Status (2013-2014)
IUCN Red List version 2014.3: Table 7 Last Updated: 13 November 2014 Table 7: Species changing IUCN Red List Status (2013-2014) Published listings of a species' status may change for a variety of reasons (genuine improvement or deterioration in status; new information being available that was not known at the time of the previous assessment; taxonomic changes; corrections to mistakes made in previous assessments, etc. To help Red List users interpret the changes between the Red List updates, a summary of species that have changed category between 2013 (IUCN Red List version 2013.2) and 2014 (IUCN Red List version 2014.3) and the reasons for these changes is provided in the table below. IUCN Red List Categories: EX - Extinct, EW - Extinct in the Wild, CR - Critically Endangered, EN - Endangered, VU - Vulnerable, LR/cd - Lower Risk/conservation dependent, NT - Near Threatened (includes LR/nt - Lower Risk/near threatened), DD - Data Deficient, LC - Least Concern (includes LR/lc - Lower Risk, least concern). Reasons for change: G - Genuine status change (genuine improvement or deterioration in the species' status); N - Non-genuine status change (i.e., status changes due to new information, improved knowledge of the criteria, incorrect data used previously, taxonomic revision, etc.); E - Previous listing was an Error. IUCN Red List IUCN Red Reason for Red List Scientific name Common name (2013) List (2014) change version Category Category MAMMALS Allocebus trichotis Hairy-eared Dwarf Lemur DD VU N 2014.1 Avahi betsileo Betsileo Woolly Lemur -

TAIWAN Formosan Endemics
TAIWAN Formosan Endemics A Tropical Birding Custom Tour June 14 – 22, 2019 Guides: Ken Behrens & Kuan Chieh Hung/貫捷洪 Report and photos by Ken Behrens TOUR SUMMARY Taiwan is a gem of a country that lies off the coast of East Asia. Although its mention evokes huge urban areas and massive populations in the minds of most, Taiwan actually boasts rugged mountains and vast wilderness areas. These support a rich array of wildlife, including dozens of endemic bird species and even more endemic subspecies, many of which are potential future splits. Taiwan is a comfortable place to travel, with modern hotels, world-class roads, and excellent food. Whether you’re an experienced Asia-phile birder seeking out the island’s endemic birds, or a newcomer to this part of the world, Taiwan is an excellent place to visit. This short trip was a bit unusual in a couple respects. For one, it was during the summer, whereas most birding tours run during the spring season. Although in June, the winter and passage birds were gone, it was if anything better for the endemic birds. This custom trip was also somewhat unusual in its hybrid approach, as we tried to see all the endemic birds, photograph as many birds as possible (as on a “Birding with a Camera” tour), plus to see as much other wildlife as we could manage. We were highly successful on all fronts. Taiwan: Formosan Endemics June 14-22, 2019 With the endemic birds, we saw all 28 endemic species recognized on the current Clements list, plus several highly likely future splits, and dozens more endemic subspecies, some of which may be elevated to full endemic species in the future. -

Near Threatened Species NEAR THREATENED SPECIES
Near Threatened species NEAR THREATENED SPECIES 2483 Threatened birds of Asia DWARF CASSOWARY Casuarius bennetti occurs in New Guinea (Papua, formerly Irian Jaya, Indonesia and Papua New Guinea) and, presumably as a long-established introduction, on New Britain, where it is a forest species occurring into the mountains and occasionally to the treeline at 3,600 m (Coates 1985, Beehler et al. 1986). Although probably tolerant of moderate habitat degradation, logging opens up previously inaccessible areas to hunters; despite heavy hunting pressure, it remains relatively common over a wide altitudinal range (Coates 1985, Beehler et al. 1986, B. M. Beehler in litt. 2000, A. Mack in litt. 1999). It is judged to have a substantial population and to be declining more slowly than the other larger and more lowland cassowaries Casuarius. Criteria nearly met: A1b,d; A2b,d. PYGMY CORMORANT Phalacrocorax pygmeus breeds in (all data for pairs, and unless otherwise indicated all information from Crivelli et al. 1996) Bulgaria (20–180), Greece (1,250– 1,310) (Kazantzidis and Nazirides 1999), Italy (30–50) (M. Passarella in litt. 1999), Moldova (30–500), Hungary, Romania (4,000–7,000), Turkey (1,000–1,500) (Eken and Magnin in press), Slovakia, Yugoslavia (1,000–1,200), F.Y.R.O. Macedonia, Croatia, Ukraine (20–320), Russia, Iran (20–30), Azerbaijan (14,749 estimated in 1986), Kazakhstan, Tajikistan, Turkmenistan, and Uzebekistan, plus, possibly, south-east Iraq, and it winters primarily in Albania, Greece, Yugoslavia, F.Y.R.O. Macedonia, Turkey, Cyprus, Iraq, Iran, Azerbaijan and also Israel, Bulgaria, Romania and Syria. There is one record from Pakistan (Grimmett et al. -
Taiwan: Formosan Endemics Set Departure Tour 14Th – 27Th April, 2014
Taiwan: Formosan Endemics Set departure tour 14th – 27th April, 2014 Tour leader: Charley Hesse Report by Charley Hesse Photos by Charley Hesse & Graham Ekins Swinhoe’s Pheasant dazzled us at the feeding station with its jewel-like plumage (Graham Ekins) This year’s Taiwan Set Departure tour was again a resounding success. Not only did we see an impressive 209 species of birds, including all the endemics (which have increased now to 24 species) and the vast majority of endemic subspecies, but thanks to the efforts of the tour participants we had the best ever lists for reptiles, amphibians and our first butterfly list. We were lucky to have fine weather for most of the trip, which made the birding comfortable and enabled us to enjoy the breath-taking scenery of this beautiful island. Again, clients were impressed at how developed, safe and friendly Taiwan is and also at the great facilities in the parks we visited. The highlights of our trip included great views of male Swinhoe’s & Mikado Pheasants and Taiwan Partridge at feeding sites, great night viewing with Formosan Serow, Reeves Munjac and Red-and- white Flying Squirrel also at Dasyueshan, finding the MEGA Himalayan or Chinese Tawny Owl at Hehuanshan and the incredibly elusive, white-headed Island Thrush at Cingjing, plus amazing butterfly viewing at Shuangliou. Tropical Birding is the only major bird tour company with an office in Taiwan, and this together with our ever-growing experience and knowledge of great stake outs for difficult species mean that we are probably the premier bird tour company here. -
Bird List Column A: Number of Tours (Out of 11) on Which This Species Was Seen
Taiwan Cumulative Bird List Column A: number of tours (out of 11) on which this species was seen. Column B: total number of days that the species was recorded on the 2018 tour Column C: maximum daily count for that particular species on the 2018 tour Column D: H = heard only; (H) = Heard more than seen E = endemic species; Essp = endemic subspecies; (I) = introduced species A B C D 1 Mandarin Duck Aix galericulata 2 Gadwall Anas strepera 8 Eurasian Wigeon 1 44 Anas penelope 2 Mallard Anas p. platyrhynchos 11 Eastern Spot-billed Duck 3 12 Anas zonorhyncha 6 Northern Shoveler Anas clypeata 2 Northern Pintail Anas acuta 3 Garganey Anas querquedula 3 Eurasian Teal Anas c. crecca 4 Tufted Duck Aythya fuligula 1 Greater Scaup Aythya marila nearctica 11 Taiwan Partridge E. 5 9 (H) Arborophila crudigularis 11 Taiwan Bamboo Partridge E. 4 4 (H) Bambusicola sonorivox 11 Swinhoe's Pheasant E. 2 4 Lophura swinhoii 10 Mikado Pheasant E. 2 2 Syrmaticus mikado 11 Common Pheasant Essp. 4 5 Phasianus colchicus formosanus 8 Streaked Shearwater 2 4 Calonectris leucomelas 1 Wedge-tailed Shearwater Puffinus pacificus 2 Streaked or Wedge-tailed Shearwater C. leucomelas or P. pacificus 1 Short-tailed Shearwater Puffinus tenuirostris 6 Sooty or Short-tailed Shearwater Puffinus griseus or P. tenuirostris 3 Shearwater sp. Puffinus sp. 4 Bulwer's Petrel Bulweria bulwerii 11 Little Grebe 2 70 Tachybaptus ruficollis philippensis 1 Greater Flamingo Phoenicopterus roseus 7 African Sacred Ibis (I.) 2 100 Threskiornis aethiopicus 1 Oriental Stork Ciconia boyciana 1 Glossy Ibis Plegadis falcinellus 4 Eurasian Spoonbill Platalea leucorodia 11 Black-faced Spoonbill 2 43 Platalea minor 10 Yellow Bittern 2 10 Ixobrychus sinensis ________________________________________________________________________________________________________ WINGS ● 1643 N. -
An Introduction to Taiwan
An introduction to the beautiful island lifeoftaiwan.com [email protected] !1 Index An Introduction to Taiwan ..................................................................................4 People of Taiwan ..............................................................................................6 Taiwan’s Languages .......................................................................................10 Geography and Climate ..................................................................................11 Climate ..........................................................................................................14 Nature and Ecology ........................................................................................15 Food and Drink ...............................................................................................18 Religion in Taiwan ...........................................................................................25 Destinations ...................................................................................................29 Taipei .............................................................................................................30 Taroko Gorge .................................................................................................35 Sun Moon Lake ..............................................................................................40 Alishan ...........................................................................................................43 Tainan -

Evidence for Wide Dispersal in a Stem Galliform Clade from a New Small-Sized Middle Eocene Pangalliform (Aves: Paraortygidae) from the Uinta Basin of Utah (USA)
diversity Article Evidence for Wide Dispersal in a Stem Galliform Clade from a New Small-Sized Middle Eocene Pangalliform (Aves: Paraortygidae) from the Uinta Basin of Utah (USA) Thomas A. Stidham 1,2,3,*, K.E. Beth Townsend 4 and Patricia A. Holroyd 5 1 Key Laboratory of Vertebrate Evolution and Human Origins of the Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China 2 CAS—Center for Excellence in Life and Paleoenvironment, Beijing 100044, China 3 University of Chinese Academy of Sciences, Beijing 10049, China 4 Department of Anatomy, College of Graduate Studies, Midwestern University, 19555 N. 59th Avenue, Glendale, AZ 85308, USA; [email protected] 5 Museum of Paleontology, University of California, Berkeley, CA 94720, USA; [email protected] * Correspondence: [email protected] Received: 16 January 2020; Accepted: 19 February 2020; Published: 28 February 2020 Abstract: A new bird coracoid from the Uinta Formation in the Uinta Basin in Utah (USA) records the presence of the only known pangalliform from the middle Eocene of North America, occurring in a >15 million year gap in their history. This fossil represents a new taxon, informally termed the Uintan paraortygid, which is also currently the best-supported record of the extinct Paraortygidae in North America (and among the oldest records of the group in the world). The specimen exhibits a derived enlarged procoracoid prominence with a small procoracoid process, and concave elliptical scapular cotyle that are shared with the middle Eocene paraortygids, Xorazmortyx and Scopelortyx; however, the Uintan paraortygid also has a possibly autapomorphic (pneumatic) fossa adjacent to the scapular cotyle. -

Taiwan Spring Birding 7Th to 19Th May 2019 (12 Days) Trip Report
Taiwan Spring Birding 7th to 19th May 2019 (12 days) Trip Report Swinhoe’s Pheasant, Dasyueshan Road by Paul Matson Tour Leaders: Richard Foster & Erik Forsyth Compiled by Tour Leader, Erik Forsyth Top 5 Birds as voted by the participants: 1) Fairy Pitta 2) White-whiskered Laughingthrush 3) Swinhoe’s Pheasant 4) Mikado Pheasant 5) Cinnamon Bittern/Pheasant-tailed Jacana Rockjumper Birding Tours View more tours to Taiwan Trip Report – RBL Taiwan - Spring Birding 2019 2 Tour Summary We began our tour of the green island in the vibrant city of Taipei, where a few walks around the botanical garden produced more than a couple of great birds, including endemic species and subspecies. Best of all were Taiwan Barbet and Taiwan Scimitar Babbler, and Light-vented and Black Bulbuls, as well as the stunning White-rumped Shama with its beautiful song. The star of the non-endemics was the Malayan Night Heron, which was very approachable. In the afternoon, we visited the Chiang Kai-shek Memorial Park, where we added Black-collared and Asian Glossy Starlings, and Javan and Common Myna’s, and then adding a further three Malayan Night Heron. The following day, we began our journey south and into the foothills around the Shihmen Reservoir. The weather held off just long enough for us to see most of our targets. First to fall was a small family flock of beautifully plumaged Taiwan Blue Magpies, which we watched at length, and shortly afterwards, the Taiwan Whistling Thrush appeared. We also recorded Red-rumped and Barn Swallows, and several Taiwan Barbets.