Willows (Salix Spp.) As Pollen and Nectar Sources for Sustaining Fruit and Berry Pollinating Insects
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Willows of Interior Alaska
1 Willows of Interior Alaska Dominique M. Collet US Fish and Wildlife Service 2004 2 Willows of Interior Alaska Acknowledgements The development of this willow guide has been made possible thanks to funding from the U.S. Fish and Wildlife Service- Yukon Flats National Wildlife Refuge - order 70181-12-M692. Funding for printing was made available through a collaborative partnership of Natural Resources, U.S. Army Alaska, Department of Defense; Pacific North- west Research Station, U.S. Forest Service, Department of Agriculture; National Park Service, and Fairbanks Fish and Wildlife Field Office, U.S. Fish and Wildlife Service, Department of the Interior; and Bonanza Creek Long Term Ecological Research Program, University of Alaska Fairbanks. The data for the distribution maps were provided by George Argus, Al Batten, Garry Davies, Rob deVelice, and Carolyn Parker. Carol Griswold, George Argus, Les Viereck and Delia Person provided much improvement to the manuscript by their careful editing and suggestions. I want to thank Delia Person, of the Yukon Flats National Wildlife Refuge, for initiating and following through with the development and printing of this guide. Most of all, I am especially grateful to Pamela Houston whose support made the writing of this guide possible. Any errors or omissions are solely the responsibility of the author. Disclaimer This publication is designed to provide accurate information on willows from interior Alaska. If expert knowledge is required, services of an experienced botanist should be sought. Contents -
Phytoremediation of a Highly Creosote- Contaminated Soil by Means of Salix Viminalis
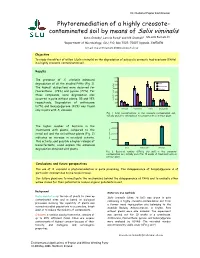
International Poplar Commission Phytoremediation of a highly creosote- contaminated soil by means of Salix viminalis Karin Önneby1, Leticia Pizzul1 and Ulf Granhall1 Mauritz Ramstedt 1Department of Microbiology, SLU, P.O. Box 7025, 75007 Uppsala, SWEDEN Email:E-mail: mauritz.ramstedt@m [email protected] Objective To study the effect of willow (Salix viminalis) on the degradation of polycyclic aromatic hydrocarbons (PAHs) in a highly creosote-contaminated soil. Results 1600 The presence of S. viminalis enhanced 1400 Initial degradation of all the studied PAHs (Fig. 1). 1200 The highest dissipations were observed for Without plant 1000 With plant fluoranthene (79%) and pyrene (77%). For 800 those compounds, some degradation also 600 occurred in pots without plants, 35 and 19% 400 Concentration (mg/kg dw) Concentration respectively. Degradation of anthracene 200 (67%) and benzo[a]pyrene (43%) was found 0 Anthracene Fluoranthene Pyrene Benzo[a]pyrene only in pots with S. viminalis. Fig. 1. PAH concentrations in the creosote-contaminated soil, initially and after 10 weeks of treatment with or without plant. 4000000 The higher number of bacteria in the 3000000 treatments with plants, compared to the initial soil and the soil without plants (Fig. 2) 2000000 indicates an increase in microbial activity. CFU (dw) /g soil 1000000 This activity, and possibly a higher release of biosurfactants, could explain the enhanced 0 degradation obtained with plants. Initial Without plant With plant Fig. 2. Bacterial number (CFU/g dry soil) in the creosote- contaminated soil, initially and after 10 weeks of treatment with or without plant. Conclusions and future perspectives The use of S. -

Chrysomela 43.10-8-04
CHRYSOMELA newsletter Dedicated to information about the Chrysomelidae Report No. 43.2 July 2004 INSIDE THIS ISSUE Fabreries in Fabreland 2- Editor’s Page St. Leon, France 2- In Memoriam—RP 3- In Memoriam—JAW 5- Remembering John Wilcox Statue of 6- Defensive Strategies of two J. H. Fabre Cassidine Larvae. in the garden 7- New Zealand Chrysomelidae of the Fabre 9- Collecting in Sholas Forests Museum, St. 10- Fun With Flea Beetle Feces Leons, France 11- Whither South African Cassidinae Research? 12- Indian Cassidinae Revisited 14- Neochlamisus—Cryptic Speciation? 16- In Memoriam—JGE 16- 17- Fabreries in Fabreland 18- The Duckett Update 18- Chrysomelidists at ESA: 2003 & 2004 Meetings 19- Recent Chrysomelid Literature 21- Email Address List 23- ICE—Phytophaga Symposium 23- Chrysomela Questionnaire See Story page 17 Research Activities and Interests Johan Stenberg (Umeå Univer- Duane McKenna (Harvard Univer- Eduard Petitpierre (Palma de sity, Sweden) Currently working on sity, USA) Currently studying phyloge- Mallorca, Spain) Interested in the cy- coevolutionary interactions between ny, ecological specialization, population togenetics, cytotaxonomy and chromo- the monophagous leaf beetles, Altica structure, and speciation in the genus somal evolution of Palearctic leaf beetles engstroemi and Galerucella tenella, and Cephaloleia. Needs Arescini and especially of chrysomelines. Would like their common host plant Filipendula Cephaloleini in ethanol, especially from to borrow or exchange specimens from ulmaria (meadow sweet) in a Swedish N. Central America and S. America. Western Palearctic areas. Archipelago. Amanda Evans (Harvard University, Maria Lourdes Chamorro-Lacayo Stefano Zoia (Milan, Italy) Inter- USA) Currently working on a phylogeny (University of Minnesota, USA) Cur- ested in Old World Eumolpinae and of Leptinotarsa to study host use evolu- rently a graduate student working on Mediterranean Chrysomelidae (except tion. -

Kenai National Wildlife Refuge Species List, Version 2018-07-24
Kenai National Wildlife Refuge Species List, version 2018-07-24 Kenai National Wildlife Refuge biology staff July 24, 2018 2 Cover image: map of 16,213 georeferenced occurrence records included in the checklist. Contents Contents 3 Introduction 5 Purpose............................................................ 5 About the list......................................................... 5 Acknowledgments....................................................... 5 Native species 7 Vertebrates .......................................................... 7 Invertebrates ......................................................... 55 Vascular Plants........................................................ 91 Bryophytes ..........................................................164 Other Plants .........................................................171 Chromista...........................................................171 Fungi .............................................................173 Protozoans ..........................................................186 Non-native species 187 Vertebrates ..........................................................187 Invertebrates .........................................................187 Vascular Plants........................................................190 Extirpated species 207 Vertebrates ..........................................................207 Vascular Plants........................................................207 Change log 211 References 213 Index 215 3 Introduction Purpose to avoid implying -

Cultivated Willows Would Not Be Appropriate Without Mention of the ‘WEEPING WILLOW’
Alexander Robertson Notes on willows cultivated in Scotland and a few sketches of willows sampled from my clonal collection HISTORICAL NOTES Since the knowledge of willows is of great antiquity, it is with the ancient Greeks and Romans we shall begin, for among these people numerous written records remain. The growth habit, ecology, cultivation and utilization of willows was well— understood by Theophrastus, Ovid, Herodotus, Pliny and Dioscorides. Virgil was also quite familiar with willow, e.g. Damoetas complains that: “Galatea, saucy girl, pelts me with apples and then runs off to the willows”. ECLOGIJE III and of foraging bees: “Far and wide they feed on arbutus, pale-green willows, on cassia and ruddy crocus .. .“ GEORGICS IV Theophrastus of Eresos (370—285 B.C.) discussed many aspects of willows throughout his Enquiry into Plants including habitats, wood quality, coppicing and a variety of uses. Willows, according to Theophrastus are lovers of wet places and marshes. But he also notes certain amphibious traits of willows growing in mountains and plains. To Theophrastus they appeared to possess no fruits and quite adequately reproduced themselves from roots, were tolerant to flooding and frequent coppicing. “Even willows grow old and when they are cut, no matter at what height, they shoot up again.” He described the wood as cold, tough, light and resilient—qualities which made it useful for a variety of purposes, especially shields. Such were the diverse virtues of willow that he suggested introducing it for plant husbandry. Theophrastus noted there were many different kinds of willows; three of the best known being black willow (Salix fragilis), white willow (S. -

Bee-Plant Networks: Structure, Dynamics and the Metacommunity Concept
Ber. d. Reinh.-Tüxen-Ges. 28, 23-40. Hannover 2016 Bee-plant networks: structure, dynamics and the metacommunity concept – Anselm Kratochwil und Sabrina Krausch, Osnabrück – Abstract Wild bees play an important role within pollinator-plant webs. The structure of such net- works is influenced by the regional species pool. After special filtering processes an actual pool will be established. According to the results of model studies these processes can be elu- cidated, especially for dry sandy grassland habitats. After restoration of specific plant com- munities (which had been developed mainly by inoculation of plant material) in a sandy area which was not or hardly populated by bees before the colonization process of bees proceeded very quickly. Foraging and nesting resources are triggering the bee species composition. Dis- persal and genetic bottlenecks seem to play a minor role. Functional aspects (e.g. number of generalists, specialists and cleptoparasites; body-size distributions) of the bee communities show that ecosystem stabilizing factors may be restored rapidly. Higher wild-bee diversity and higher numbers of specialized species were found at drier plots, e.g. communities of Koelerio-Corynephoretea and Festuco-Brometea. Bee-plant webs are highly complex systems and combine elements of nestedness, modularization and gradients. Beside structural com- plexity bee-plant networks can be characterized as dynamic systems. This is shown by using the metacommunity concept. Zusammenfassung: Wildbienen-Pflanzenarten-Netzwerke: Struktur, Dynamik und das Metacommunity-Konzept. Wildbienen spielen eine wichtige Rolle innerhalb von Bestäuber-Pflanzen-Netzwerken. Ihre Struktur wird vom jeweiligen regionalen Artenpool bestimmt. Nach spezifischen Filter- prozessen bildet sich ein aktueller Artenpool. -

List of Publications —MARIA GREGER
List of Publications —MARIA GREGER Scientific Publications (Paper with review system = R; Included in thesis = X) 1. RX Greger M. & Lindberg S., 1986. Effects of Cd2+ and EDTA on young sugar beets (Beta vulgaris). I. Cd2+uptake and sugar accumulation. — Physiologia Plantarum 66: 69-74. 2. RX Greger M. & Lindberg S., 1987. Effects of Cd2+ and EDTA on young sugar beets (Beta vulgaris). II. Net uptake and distribution of Mg2+, Ca2+ and Fe2+/Fe3+. — Physiologia Plantarum 69: 81-86. 3. RX Greger M., 1989. Cadmium Effects on Carbohydrate Metabolism in Sugar Beet (Beta vulgaris). — Thesis, Stockholm University, ISBN 91-7146-717-3 4. R Kronestedt-Robards E. C., Greger M. & Robards A. W., 1989. The nectar of the Strelizia Reginæ flower. — Physiologia Plantarum 77: 341-346. 5. R X Greger M., Brammer E. S., Lindberg S., Larsson G. & Idestam-Almquist J., 1991. Uptake and physiological effects of cadmium in sugar beets (Beta vulgaris) related to mineral provision. — Journal of Experimental Botany 42: 729-737. 6. R Lindberg S., Szynkier K. & Greger M., 1991. Aluminum effects on transmembrane potential in cells of fibrous roots of sugar beet. — Physiologia Plantarum 83: 54-62. 7. R X Greger M. & Ögren E., 1991. Direct and indirect effects of Cd2+ on the photosynthesis and CO2-assimilation in sugar beets (Beta vulgaris). — Physiologia Plantarum 83: 129-135. 8. R Greger M. & Kautsky L., 1991. Effects of Cu, Pb, and Zn on two species of Potamogeton grown in field conditions. — Vegetatio 97: 173-184. 9. R X Greger M. & Bertell G., 1992. Effects of Ca2+ and Cd2+ on the carbohydrate metabolism in sugar beet (Beta vulgaris). -

CONTAINER WILLOWS in the Southwestern US Using Seeds
(Taken from Native Plant Journal, vol. 4 | no. 2 | fall 2003) PROPAGATION PROTOCOL FOR CONTAINER WILLOWS In the Southwestern US using Seeds David R Dreesen | KEY WORDS Salix arizonica, S. bebbiana, The genetic and sexual diversity provided by seed S. exigua, S. irrorata, S. propagation of Salix L. (Salicaceae) species is an important scouleriana, arizona willow, consideration when planning riparian restoration projects. A bebb's willow, coyote willow, standard seed propagation protocol has been successful with bluestern willow, Scouler willow, a number of Salix species from montane habitats as well as height growth, stem diameter low elevation floodplains. Despite the very small seed size, growth, caliper, Salicaceae, most Salix are extremely rapid growers, which helps negate controlled release fertilizer some of the advantage typically attributed to cutting NOMENCLATURE ITIS (2002) propagation. Perhaps the largest hurdle to seedling propagation of Salix is the collection of seeds in the wild. However, rapid development of nursery seed orchards is possible using reproductive cuttings representing a mix of male and female plants or seedling stock which rapidly becomes reproductive for many species. Two principal differences between Salix and Populus L. (Salicaceae) influence seedling propagation strategies if nursery seed stock plants are desired. The first relates to the location of juvenile and mature wood in the 2 genera and where cuttings are usually taken. Often, Populus cuttings are juvenile because they are taken from stems close to the root crown, not from mature stems in the upper canopy. Therefore, cuttings are unlikely to have preformed floral buds that can be forced in the current year. -

Phytoremediation of Metal Contamination Using Salix (Willows)
University of Denver Digital Commons @ DU Electronic Theses and Dissertations Graduate Studies 1-1-2015 Phytoremediation of Metal Contamination Using Salix (Willows) Gordon J. Kersten University of Denver Follow this and additional works at: https://digitalcommons.du.edu/etd Part of the Ecology and Evolutionary Biology Commons, and the Other Environmental Sciences Commons Recommended Citation Kersten, Gordon J., "Phytoremediation of Metal Contamination Using Salix (Willows)" (2015). Electronic Theses and Dissertations. 1034. https://digitalcommons.du.edu/etd/1034 This Thesis is brought to you for free and open access by the Graduate Studies at Digital Commons @ DU. It has been accepted for inclusion in Electronic Theses and Dissertations by an authorized administrator of Digital Commons @ DU. For more information, please contact [email protected],[email protected]. Phytoremediation of Metal Contamination using Salix (willows) ______________________ A Thesis Presented to The Faculty of Natural Sciences and Mathematics University of Denver ______________________ In Partial Fulfillment Of the Requirements for the Degree Master of Science ______________________ By Gordon J. Kersten August 2015 Advisor: Martin F. Quigley Author: Gordon J. Kersten Title: Phytoremediation of Metal Contamination using Salix (willows) Advisor: Martin F. Quigley Degree Date: August 2015 ABSTRACT Abandoned hardrock mines and the resulting Acid Mine Drainage (AMD) are a source of vast, environmental degradation that are toxic threats to plants, animals, and humans. Cadmium (Cd) and lead (Pb) are metal contaminants often found in AMD. In my mine outwash water samples, cadmium and lead concentrations were 19 and 160 times greater than concentrations in control waterways, and 300 and 40 times greater than EPA Aquatic Life Use water quality standards, respectively. -

Guide to the Willows of Shoshone National Forest
United States Department of Agriculture Guide to the Willows Forest Service Rocky Mountain Research Station of Shoshone National General Technical Report RMRS-GTR-83 Forest October 2001 Walter Fertig Stuart Markow Natural Resources Conservation Service Cody Conservation District Abstract Fertig, Walter; Markow, Stuart. 2001. Guide to the willows of Shoshone National Forest. Gen. Tech. Rep. RMRS-GTR-83. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 79 p. Correct identification of willow species is an important part of land management. This guide describes the 29 willows that are known to occur on the Shoshone National Forest, Wyoming. Keys to pistillate catkins and leaf morphology are included with illustrations and plant descriptions. Key words: Salix, willows, Shoshone National Forest, identification The Authors Walter Fertig has been Heritage Botanist with the University of Wyoming’s Natural Diversity Database (WYNDD) since 1992. He has conducted rare plant surveys and natural areas inventories throughout Wyoming, with an emphasis on the desert basins of southwest Wyoming and the montane and alpine regions of the Wind River and Absaroka ranges. Fertig is the author of the Wyoming Rare Plant Field Guide, and has written over 100 technical reports on rare plants of the State. Stuart Markow received his Masters Degree in botany from the University of Wyoming in 1993 for his floristic survey of the Targhee National Forest in Idaho and Wyoming. He is currently a Botanical Consultant with a research emphasis on the montane flora of the Greater Yellowstone area and the taxonomy of grasses. Acknowledgments Sincere thanks are extended to Kent Houston and Dave Henry of the Shoshone National Forest for providing Forest Service funding for this project. -

Towards Sustainable Crop Pollination Services Measures at Field, Farm and Landscape Scales
EXTENSION OF KNOWLEDGE BASE ADAPTIVE MANAGEMENT CAPACITY BUILDING MAINSTREAMING TOWARDS SUSTAINABLE CROP POLLINATION SERVICES MEASURES AT FIELD, FARM AND LANDSCAPE SCALES POLLINATION SERVICES FOR SUSTAINABLE AGRICULTURE POLLINATION SERVICES FOR SUSTAINABLE AGRICULTURE TOWARDS SUSTAINABLE CROP POLLINATION SERVICES MEASURES AT FIELD, FARM AND LANDSCAPE SCALES B. Gemmill-Herren, N. Azzu, A. Bicksler, and A. Guidotti [eds.] FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS ROME, 2020 Required citation: FAO. 2020. Towards sustainable crop pollination services – Measures at field, farm and landscape scales. Rome. https://doi.org/10.4060/ca8965en The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations (FAO) concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of specific companies or products of manufacturers, whether or not these have been patented, does not imply that these have been endorsed or recommended by FAO in preference to others of a similar nature that are not mentioned. The views expressed in this information product are those of the author(s) and do not necessarily reflect the views or policies of FAO. ISBN 978-92-5-132578-0 © FAO, 2020 Some rights reserved. This work is made available under the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 IGO licence (CC BY-NC-SA 3.0 IGO; https://creativecommons.org/licenses/by-nc-sa/3.0/igo/legalcode). Under the terms of this licence, this work may be copied, redistributed and adapted for non-commercial purposes, provided that the work is appropriately cited. -

Clarkella (PDF)
Fl. China 19: 89–90. 2011. 17. CLARKELLA J. D. Hooker, Fl. Brit. India 3: 46. 1880. 岩上珠属 yan shang zhu shu Chen Tao (陈涛); Charlotte M. Taylor Small herbs from tubers, apparently perennial, unarmed, usually with a cluster of prophylls at stem base. Raphides present. Leaves opposite, isophyllous or sometimes markedly anisophyllous at basalmost node, without domatia; stipules persistent, inter- petiolar and fused to petioles, triangular, perhaps sometimes glandular on margins. Inflorescences terminal, cymose, umbelliform, or subfasciculate, few to several flowered, pedunculate or sessile and tripartite, bracteate. Flowers pedicellate, bisexual with biology unknown. Calyx limb well developed, markedly reticulate veined, 5(–7)-lobed for ca. 1/2 length. Corolla white, slenderly salverform or funnelform-salverform, glabrous inside; lobes 5, valvate in bud. Stamens 5, inserted at base of corolla tube, included; filaments short. Ovary 2-celled, ovules numerous in each cell on axile placentas inserted not far below middle of septum; disk pilosulous; stig- ma 2-lobed, pubescent. Fruit indehiscent, obconical, dry, with funnelform calyx limb persistent; seeds numerous, small, subellipsoid, black papillose or -granular. One species: China, N India, N Myanmar, Thailand. This is a poorly known species or genus that apparently prefers wet limestone substrates and is similar to Pseudopyxis. 1. Clarkella nana (Edgeworth) J. D. Hooker, Fl. Brit. India 3: apex acute to rounded; secondary veins 4–8 pairs; stipules tri- 46. 1880. angular to narrowly triangular, 0.2–0.5 mm. Inflorescence 0.8–3 cm, 3–25-flowered, villosulous to puberulent; bracts ovate 岩上珠 yan shang zhu to narrowly oblong, 0.5–12 mm; pedicels 1–3.5 mm.