Cerebral Cortex 595
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cerebral Cortex Structure, Function, Dysfunction Reading Ch 10 Waxman Dental Neuroanatomy Lecture Suzanne Stensaas, Ph.D
Cerebral Cortex Structure, Function, Dysfunction Reading Ch 10 Waxman Dental Neuroanatomy Lecture Suzanne Stensaas, Ph.D. March 15, 2011 Anatomy Review • Lobes and layers • Brodmann’s areas • Vascular Supply • Major Neurological Findings – Frontal, Parietal, Temporal, Occipital, Limbic • Quiz Questions? Types of Cortex • Sensory • Motor • Unimodal association • Multimodal association necessary for language, reason, plan, imagine, create Structure of neocortex (6 layers) The general pattern of primary, association and mulimodal association cortex (Mesulam) Brodmann, Lateral Left Hemisphere MCA left hemisphere from D.Haines ACA and PCA -Haines Issues of Functional Localization • Earliest studies -Signs, symptoms and note location • Electrical discharge (epilepsy) suggested function • Ablation - deficit suggest function • Reappearance of infant functions suggest loss of inhibition (disinhibition), i.e. grasp, suck, Babinski • Variabilities in case reports • Linked networks of afferent and efferent neurons in several regions working to accomplish a task • Functional imaging does not always equate with abnormal function associated with location of lesion • fMRI activation of several cortical regions • Same sign from lesions in different areas – i.e.paraphasias • Notion of the right hemisphere as "emotional" in contrast to the left one as "logical" has no basis in fact. Limbic System (not a true lobe) involves with cingulate gyrus and the • Hippocampus- short term memory • Amygdala- fear, agression, mating • Fornix pathway to hypothalamus • -

Combined Structural and Diffusion Tensor Imaging Detection of Ischemic Injury in Moyamoya Disease: Relation to Disease Advancement and Cerebral Hypoperfusion
CLINICAL ARTICLE Combined structural and diffusion tensor imaging detection of ischemic injury in moyamoya disease: relation to disease advancement and cerebral hypoperfusion Ken Kazumata, MD, PhD,1 Kikutaro Tokairin, MD,1 Masaki Ito, MD, PhD,1 Haruto Uchino, MD, PhD,1 Taku Sugiyama, MD, PhD,1 Masahito Kawabori, MD, PhD,1 Toshiya Osanai, MD, PhD,1 Khin Khin Tha, MD, PhD,2 and Kiyohiro Houkin, MD, PhD1 1Department of Neurosurgery, Hokkaido University Graduate School of Medicine; and 2Clinical Research and Medical Innovation Center, Hokkaido University Hospital, Sapporo, Japan OBJECTIVE The microstructural integrity of gray and white matter is decreased in adult moyamoya disease, suggesting covert ischemic injury as a mechanism of cognitive dysfunction. Establishing a microstructural brain imaging marker is critical for monitoring cognitive outcomes following surgical interventions. The authors of the present study determined the pathophysiological basis of altered microstructural brain injury in relation to advanced arterial occlusion, cerebral hypoperfusion, and cognitive function. METHODS The authors examined 58 patients without apparent brain lesions and 30 healthy controls by using structural MRI, as well as diffusion tensor imaging (DTI). Arterial occlusion in each hemisphere was classified as early or ad- vanced stage based on MRA and posterior cerebral artery (PCA) involvement. Regional cerebral blood flow (rCBF) was measured with N-isopropyl-p-[123I]-iodoamphetamine SPECT. Furthermore, cognitive performance was examined using the Wechsler Adult Intelligence Scale, Third Edition and the Trail Making Test (TMT). Both voxel- and region of inter- est–based analyses were performed for groupwise comparisons, as well as correlation analysis, using parameters such as cognitive test scores; gray matter volume; fractional anisotropy (FA) of association fiber tracts, including the inferior frontooccipital fasciculus (IFOF) and superior longitudinal fasciculus (SLF); PCA involvement; and rCBF. -

Network Architecture of the Cerebral Nuclei (Basal Ganglia) Association and Commissural Connectome
Network architecture of the cerebral nuclei (basal ganglia) association and commissural connectome Larry W. Swansona,1, Olaf Spornsb, and Joel D. Hahna aDepartment of Biological Sciences, University of Southern California, Los Angeles, CA 90089; and bDepartment of Psychological and Brain Sciences, Indiana University, Bloomington, IN 47405 Contributed by Larry W. Swanson, August 10, 2016 (sent for review July 18, 2016; reviewed by Ann M. Graybiel and Liqun Luo) The cerebral nuclei form the ventral division of the cerebral hemi- same basic strategy and methodology applied to the rat cerebral sphere and are thought to play an important role in neural systems cortical association macroconnectome (10) but with additional an- controlling somatic movement and motivation. Network analysis alytical approaches and curation tools. In this approach a macro- was used to define global architectural features of intrinsic cerebral connection is defined as a monosynaptic axonal (directed, from/to) nuclei circuitry in one hemisphere (association connections) and connection between two nervous system gray matter regions or between hemispheres (commissural connections). The analysis was between a gray matter region and another part of the body (such as based on more than 4,000 reports of histologically defined axonal a muscle) (11, 12). All 45 gray matter regions of the cerebral nuclei connections involving all 45 gray matter regions of the rat cerebral on each side of the brain were included in the analysis. The goal of nuclei and revealed the existence of four asymmetrically intercon- this analysis was to provide global, high-level, design principles of nected modules. The modules form four topographically distinct intrinsic cerebral nuclei circuitry as a framework for more detailed research at the meso-, micro-, and nanolevels of analysis (13). -

Structure and Function of Visual Area MT
AR245-NE28-07 ARI 16 March 2005 1:3 V I E E W R S First published online as a Review in Advance on March 17, 2005 I E N C N A D V A Structure and Function of Visual Area MT Richard T. Born1 and David C. Bradley2 1Department of Neurobiology, Harvard Medical School, Boston, Massachusetts 02115-5701; email: [email protected] 2Department of Psychology, University of Chicago, Chicago, Illinois 60637; email: [email protected] Annu. Rev. Neurosci. Key Words 2005. 28:157–89 extrastriate, motion perception, center-surround antagonism, doi: 10.1146/ magnocellular, structure-from-motion, aperture problem by HARVARD COLLEGE on 04/14/05. For personal use only. annurev.neuro.26.041002.131052 Copyright c 2005 by Abstract Annual Reviews. All rights reserved The small visual area known as MT or V5 has played a major role in 0147-006X/05/0721- our understanding of the primate cerebral cortex. This area has been 0157$20.00 historically important in the concept of cortical processing streams and the idea that different visual areas constitute highly specialized Annu. Rev. Neurosci. 0.0:${article.fPage}-${article.lPage}. Downloaded from arjournals.annualreviews.org representations of visual information. MT has also proven to be a fer- tile culture dish—full of direction- and disparity-selective neurons— exploited by many labs to study the neural circuits underlying com- putations of motion and depth and to examine the relationship be- tween neural activity and perception. Here we attempt a synthetic overview of the rich literature on MT with the goal of answering the question, What does MT do? www.annualreviews.org · Structure and Function of Area MT 157 AR245-NE28-07 ARI 16 March 2005 1:3 pathway. -

Quantitative Analysis of Axon Collaterals of Single Pyramidal Cells
Yang et al. BMC Neurosci (2017) 18:25 DOI 10.1186/s12868-017-0342-7 BMC Neuroscience RESEARCH ARTICLE Open Access Quantitative analysis of axon collaterals of single pyramidal cells of the anterior piriform cortex of the guinea pig Junli Yang1,2*, Gerhard Litscher1,3* , Zhongren Sun1*, Qiang Tang1, Kiyoshi Kishi2, Satoko Oda2, Masaaki Takayanagi2, Zemin Sheng1,4, Yang Liu1, Wenhai Guo1, Ting Zhang1, Lu Wang1,3, Ingrid Gaischek3, Daniela Litscher3, Irmgard Th. Lippe5 and Masaru Kuroda2 Abstract Background: The role of the piriform cortex (PC) in olfactory information processing remains largely unknown. The anterior part of the piriform cortex (APC) has been the focus of cortical-level studies of olfactory coding, and asso- ciative processes have attracted considerable attention as an important part in odor discrimination and olfactory information processing. Associational connections of pyramidal cells in the guinea pig APC were studied by direct visualization of axons stained and quantitatively analyzed by intracellular biocytin injection in vivo. Results: The observations illustrated that axon collaterals of the individual cells were widely and spatially distrib- uted within the PC, and sometimes also showed a long associational projection to the olfactory bulb (OB). The data showed that long associational axons were both rostrally and caudally directed throughout the PC, and the intrinsic associational fibers of pyramidal cells in the APC are omnidirectional connections in the PC. Within the PC, associa- tional axons typically followed rather linear trajectories and irregular bouton distributions. Quantitative data of the axon collaterals of two pyramidal cells in the APC showed that the average length of axonal collaterals was 101 mm, out of which 79 mm (78% of total length) were distributed in the PC. -

Basic Brain Anatomy
Chapter 2 Basic Brain Anatomy Where this icon appears, visit The Brain http://go.jblearning.com/ManascoCWS to view the corresponding video. The average weight of an adult human brain is about 3 pounds. That is about the weight of a single small To understand how a part of the brain is disordered by cantaloupe or six grapefruits. If a human brain was damage or disease, speech-language pathologists must placed on a tray, it would look like a pretty unim- first know a few facts about the anatomy of the brain pressive mass of gray lumpy tissue (Luria, 1973). In in general and how a normal and healthy brain func- fact, for most of history the brain was thought to be tions. Readers can use the anatomy presented here as an utterly useless piece of flesh housed in the skull. a reference, review, and jumping off point to under- The Egyptians believed that the heart was the seat standing the consequences of damage to the structures of human intelligence, and as such, the brain was discussed. This chapter begins with the big picture promptly removed during mummification. In his and works down into the specifics of brain anatomy. essay On Sleep and Sleeplessness, Aristotle argued that the brain is a complex cooling mechanism for our bodies that works primarily to help cool and The Central Nervous condense water vapors rising in our bodies (Aristo- tle, republished 2011). He also established a strong System argument in this same essay for why infants should not drink wine. The basis for this argument was that The nervous system is divided into two major sec- infants already have Central nervous tions: the central nervous system and the peripheral too much moisture system The brain and nervous system. -
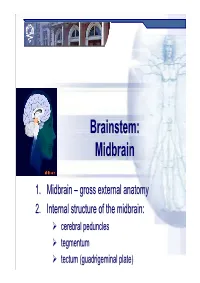
Brainstem: Midbrainmidbrain
Brainstem:Brainstem: MidbrainMidbrain 1.1. MidbrainMidbrain –– grossgross externalexternal anatomyanatomy 2.2. InternalInternal structurestructure ofof thethe midbrain:midbrain: cerebral peduncles tegmentum tectum (guadrigeminal plate) Midbrain MidbrainMidbrain –– generalgeneral featuresfeatures location – between forebrain and hindbrain the smallest region of the brainstem – 6-7g the shortest brainstem segment ~ 2 cm long least differentiated brainstem division human midbrain is archipallian – shared general architecture with the most ancient of vertebrates embryonic origin – mesencephalon main functions:functions a sort of relay station for sound and visual information serves as a nerve pathway of the cerebral hemispheres controls the eye movement involved in control of body movement Prof. Dr. Nikolai Lazarov 2 Midbrain MidbrainMidbrain –– grossgross anatomyanatomy dorsal part – tectum (quadrigeminal plate): superior colliculi inferior colliculi cerebral aqueduct ventral part – cerebral peduncles:peduncles dorsal – tegmentum (central part) ventral – cerebral crus substantia nigra Prof. Dr. Nikolai Lazarov 3 Midbrain CerebralCerebral cruscrus –– internalinternal structurestructure CerebralCerebral peduncle:peduncle: crus cerebri tegmentum mesencephali substantia nigra two thick semilunar white matter bundles composition – somatotopically arranged motor tracts: corticospinal } pyramidal tracts – medial ⅔ corticobulbar corticopontine fibers: frontopontine tracts – medially temporopontine tracts – laterally -

01 05 Lateral Surface of the Brain-NOTES.Pdf
Lateral Surface of the Brain Medical Neuroscience | Tutorial Notes Lateral Surface of the Brain 1 MAP TO NEUROSCIENCE CORE CONCEPTS NCC1. The brain is the body's most complex organ. LEARNING OBJECTIVES After study of the assigned learning materials, the student will: 1. Demonstrate the four paired lobes of the cerebral cortex and describe the boundaries of each. 2. Sketch the major features of each cerebral lobe, as seen from the lateral view, identifying major gyri and sulci that characterize each lobe. NARRATIVE by Leonard E. WHITE and Nell B. CANT Duke Institute for Brain Sciences Department of Neurobiology Duke University School of Medicine Overview When you view the lateral aspect of a human brain specimen (see Figures A3A and A102), three structures are usually visible: the cerebral hemispheres, the cerebellum, and part of the brainstem (although the brainstem is not visible in the specimen photographed in lateral view for Fig. 1 below). The spinal cord has usually been severed (but we’ll consider the spinal cord later), and the rest of the subdivisions are hidden from lateral view by the hemispheres. The diencephalon and the rest of the brainstem are visible on the medial surface of a brain that has been cut in the midsagittal plane. Parts of all of the subdivisions are also visible from the ventral surface of the whole brain. Over the next several tutorials, you will find video demonstrations (from the brain anatomy lab) and photographs (in the tutorial notes) of these brain surfaces, and sufficient detail in the narrative to appreciate the overall organization of the parts of the brain that are visible from each perspective. -

Brain Maps – the Sensory Homunculus
Brain Maps – The Sensory Homunculus Our brains are maps. This mapping results from the way connections in the brain are ordered and arranged. The ordering of neural pathways between different parts of the brain and those going to and from our muscles and sensory organs produces specific patterns on the brain surface. The patterns on the brain surface can be seen at various levels of organization. At the most general level, areas that control motor functions (muscle movement) map to the front-most areas of the cerebral cortex while areas that receive and process sensory information are more towards the back of the brain (Figure 1). Motor Areas Primary somatosensory area Primary visual area Sensory Areas Primary auditory area Figure 1. A diagram of the left side of the human cerebral cortex. The image on the left shows the major division between motor functions in the front part of the brain and sensory functions in the rear part of the brain. The image on the right further subdivides the sensory regions to show regions that receive input from somatosensory, auditory, and visual receptors. We then can divide these general maps of motor and sensory areas into regions with more specific functions. For example, the part of the cerebral cortex that receives visual input from the retina is in the very back of the brain (occipital lobe), auditory information from the ears comes to the side of the brain (temporal lobe), and sensory information from the skin is sent to the top of the brain (parietal lobe). But, we’re not done mapping the brain. -

The Human Brain Hemisphere Controls the Left Side of the Body and the Left What Makes the Human Brain Unique Is Its Size
About the brain Cerebrum (also known as the The brain is made up of around 100 billion nerve cells - each one cerebral cortex or forebrain) is connected to another 10,000. This means that, in total, we The cerebrum is the largest part of the brain. It is split in to two have around 1,000 trillion connections in our brains. (This would ‘halves’ of roughly equal size called hemispheres. The two be written as 1,000,000,000,000,000). These are ultimately hemispheres, the left and right, are joined together by a bundle responsible for who we are. Our brains control the decisions we of nerve fibres called the corpus callosum. The right make, the way we learn, move, and how we feel. The human brain hemisphere controls the left side of the body and the left What makes the human brain unique is its size. Our brains have a hemisphere controls the right side of the body. The cerebrum is larger cerebral cortex, or cerebrum, relative to the rest of the The human brain is the centre of our nervous further divided in to four lobes: frontal, parietal, occipital, and brain than any other animal. (See the Cerebrum section of this temporal, which have different functions. system. It is the most complex organ in our fact sheet for further information.) This enables us to have abilities The frontal lobe body and is responsible for everything we do - such as complex language, problem-solving and self-control. The frontal lobe is located at the front of the brain. -

Function of Cerebral Cortex
FUNCTION OF CEREBRAL CORTEX Course: Neuropsychology CC-6 (M.A PSYCHOLOGY SEM II); Unit I By Dr. Priyanka Kumari Assistant Professor Institute of Psychological Research and Service Patna University Contact No.7654991023; E-mail- [email protected] The cerebral cortex—the thin outer covering of the brain-is the part of the brain responsible for our ability to reason, plan, remember, and imagine. Cerebral Cortex accounts for our impressive capacity to process and transform information. The cerebral cortex is only about one-eighth of an inch thick, but it contains billions of neurons, each connected to thousands of others. The predominance of cell bodies gives the cortex a brownish gray colour. Because of its appearance, the cortex is often referred to as gray matter. Beneath the cortex are myelin-sheathed axons connecting the neurons of the cortex with those of other parts of the brain. The large concentrations of myelin make this tissue look whitish and opaque, and hence it is often referred to as white matter. The cortex is divided into two nearly symmetrical halves, the cerebral hemispheres . Thus, many of the structures of the cerebral cortex appear in both the left and right cerebral hemispheres. The two hemispheres appear to be somewhat specialized in the functions they perform. The cerebral hemispheres are folded into many ridges and grooves, which greatly increase their surface area. Each hemisphere is usually described, on the basis of the largest of these grooves or fissures, as being divided into four distinct regions or lobes. The four lobes are: • Frontal, • Parietal, • Occipital, and • Temporal. -

A Fiber, 9, 10, 66, 67 Abdomen, 221 Visceral Afferent, 222 Absolute
INDEX A fiber, 9, 10, 66, 67 all-or-none law, 62 Abdomen, 221 current during propagation, 62, 63 visceral afferent, 222 current loop, 64 Absolute temperature, 26 definition, 40 Acceleration depolarization phase, 38 angular, 183 duration, 38 linear, 183, 184 effect on contraction, 144 negative, 184 frequency, 50 positive, 184 generation, 88, 89 Accommodation, excitability, 60 inactivation, 48 Acetic acid, 74, 78 inhibition, 96 Acetylenoline ion current, 42, 43 cycle, 78 ion shift, 40, 42, 43 end plate, 74, 75 kinetics, 44-52 fate, 77, 78 mechanism of propagation, 62 intestinal muscle, 237 membrane conductance, 49 membrane receptor, 77-79 muscle, 129 muscarinergic transmission, 223, 225 overshoot, 38 nicotinergic transmission, 223, 225 peak, 38 quanta, 82 phase, 38 receptor, 78, 79 potassium conductance, 41 Renshaw cell, 100 propagation, 61-68 smooth muscle, 230, 231 refractory period, 50 transmitter function, 100, 101 refractory phase, 49, 50 Acetylcholinesterase, 101 repolarization, 38 ACh, 74, 75, s.a. acetylcholine rising phase, 38 Acid, fatty, 225 saltatory conduction, 64-66 Actin, 131-133, 139, 147 smooth muscle, 230, 231 Actinomycin, '312 sodium conductance, 41, 42 Action potential, 37-43 sodium deficiency, 43 Action potential tetrodotoxin, 52 after-potential, 39 threshold, 39 327 328 Index Action potential (cont.) Anion, 21 time course, 37, 38 Anococcygeal muscle, 233 trigger, 39 Anoce,58 triphasic current, 64 Antagonist inhibition, 109, 212 upstroke, 38 Anterior pens, micturition center, 241 velocity of conduction, 61 Anticholinergic substance, 313 Active transport Antidiuretic hormone, 259 membrane, 32 Aphagia, 264 sodium, 35 Aphasia Activity clock, 288 motor, 305 Adaptation, hormonal, 259 sensory, 305 Adenohypophysis Apoplexy, 196, 310 feedback system, 259 ARAS,295 hormone control, 257-259 Areflexia, 170 hypothalamus, 254 Arousal, 295 Adenosine triphosphate, 132-134, 147, 226 Arterial pressure, 247, s.a.