Report (4.723Mb)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Demobilizing and Integrating the Nicaraguan Resistance 1990-1997
The International Commission for Support and Verification Commission (CIAV) Demobilizing and Integrating the Nicaraguan Resistance 1990-1997 ii Acknowledgements: This paper is a summary English version, written by Fernando Arocena, a consultant to CIAV-OAS, based on the original Spanish report: “La Comisión Internacional de Apoyo y Verificación, La Desmovilización y Reinserción de la Resistencia Nicaragüense 1990 – 1997”, prepared by Héctor Vanolli, Diógenes Ruiz and Arturo Wallace, also consultants to the CIAV-OAS. Bruce Rickerson, Senior Specialist at the UPD revised and edited the English text. This is a publication of the General Secretariat of the Organization of American States. The ideas, thoughts, and opinions expressed are not necessarily those of the OAS or its member states. The opinions expressed are the responsibility of the authors. Correspondence should be directed to the UPD, 1889 "F" Street, N.W., 8th Floor, Washington, DC, 20006, USA. Copyright ©1998 by OAS. All rights reserved. This publication may be reproduced provided credit is given to the source. TABLE OF CONTENTS ACRONYMS................................................................................................................................ix READER'S GUIDE ..................................................................................................................... xi INTRODUCTION......................................................................................................................xiii EXECUTIVE SUMMARY ....................................................................................................... -

Baseline Study Report
Baseline Study Report MESA II Project - Better Education and Health Agreement: FFE-524-2017/025-00 Final Evaluation Report Coordinated by Project Concern International (PCI) Nicaragua August/Sept. 2017 Submitted to USDA/FAS Project “Mejor Educación y Salud (MESA)” - Nicaragua Agreement: FFE-524-2013-042-00 Submitted to: USDA/FAS Vanessa Castro, José Ramón Laguna, Patricia Callejas with collaboration from Micaela Gómez Managua, December 2017 June 4, 2019 Managua, Nicaragua i Acknowledgements The consultant team appreciates PCI Nicaragua for entrusting Asociación Nicaragua Lee with the completion of this study. In particular, we would like to acknowledge the valuable support provided by María Ángeles Argüello and María Zepeda at PCI Nicaragua-, and by officials from the Ministry of Education (MINED) in Managua and in the departmental delegations of Jinotega and the Southern Caribbean Coast Autonomous Region (RACCS). We also recognize the support given by the officials at the MINED offices in the 11 municipalities participating in the study: Jinotega, La Concordia, San Sebastian de Yali, Santa Maria de Pantasma, Bluefields, Kukra Hill, La Cruz del Río Grande, Laguna de Perlas, Desembocadura Río Grande, El Tortuguero and Corn Island. In particular, we would like to acknowledge the enthusiasm showed by the educational advisors from the aforementioned MINED municipal offices, in the administration of the instruments Our greatest gratitude and consideration to the actors of this study, the fourth-grade students from the elementary schools included in the sample, who agreed and participated with great enthusiasm. We would also like to thank the third-grade teachers who contributed by answering the questionnaire. We should also mention and thank the team of supervisors, applicators and data entry personnel, who put much dedication and effort into the collection and processing of the Early Grade Reading Assessment (EGRA) instruments, the questionnaires, and the school and classroom environment observation sheet. -

Nicaragua Expansion and Strengthening Of
PUBLIC SIMULTANEOUS DISCLOSURE DOCUMENT OF THE INTER-AMERICAN DEVELOPMENT BANK NICARAGUA EXPANSION AND STRENGTHENING OF NICARAGUA’S ELECTRICITY TRANSMISSION SYSTEM (NI-L1091) LOAN PROPOSAL This document was prepared by the project team consisting of: Héctor Baldivieso (ENE/CNI), Project Team Leader; Arnaldo Vieira de Carvalho (INE/ENE), Project Team Co-leader; Alberto Levy (INE/ENE); Carlos Trujillo (INE/ENE); Carlos Hinestrosa (INE/ENE); Stephanie Suber (INE/ENE); Juan Carlos Lazo (FMP/CNI); Santiago Castillo (FMP/CNI); María Cristina Landázuri (LEG/SGO); Denis Corrales (VPS/ESG); and Alma Reyna Selva (CID/CNI). This document is being released to the public and distributed to the Bank’s Board of Executive Directors simultaneously. This document has not been approved by the Board. Should the Board approve the document with amendments, a revised version will be made available to the public, thus superseding and replacing the original version. CONTENTS PROJECT SUMMARY I. DESCRIPTION AND RESULTS MONITORING ................................................................ 1 A. Background, problem to be addressed, and rationale ................................... 1 B. Objectives, components, and cost ................................................................ 8 C. Key results indicators ................................................................................. 10 II. FINANCING STRUCTURE AND MAIN RISKS ............................................................... 10 A. Financing instruments ............................................................................... -
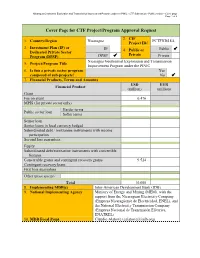
Cover Page for CTF Project/Program Approval Request
Nicaragua Geothermal Exploration and Transmission Improvement Program under the PINIC - CTF Submission - Public version – Cover page Page 1 of 6 Cover Page for CTF Project/Program Approval Request 2. CIF 1. Country/Region Nicaragua PCTFNI618A Project ID# 3. Investment Plan (IP) or IP 4. Public or Public Dedicated Private Sector Private Program (DPSP) DPSP Private Nicaragua Geothermal Exploration and Transmission 5. Project/Program Title Improvement Program under the PINIC 6. Is this a private sector program Yes composed of sub-projects? No 7. Financial Products, Terms and Amounts USD EUR Financial Product (million) (million) Grant Fee on grant 0.476 MPIS (for private sector only) Harder terms Public sector loan Softer terms Senior loan Senior loans in local currency hedged Subordinated debt / mezzanine instruments with income participation Second loss guarantees Equity Subordinated debt/mezzanine instruments with convertible features Convertible grants and contingent recovery grants 9.524 Contingent recovery loans First loss guarantees Other (please specify) Total 10.000 8. Implementing MDB(s) Inter-American Development Bank (IDB) 9. National Implementing Agency Ministry of Energy and Mining (MEM), with the support from the Nicaraguan Electricity Company (Empresa Nicaragüense de Electricidad, ENEL), and the National Electricity Transmission Company (Empresa Nacional de Transmisión Eléctrica, ENATREL) 10. MDB Focal Point Claudio Alatorre ([email protected]) Nicaragua Geothermal Exploration and Transmission Improvement Program under the PINIC - CTF Submission - Public version – Cover page Page 2 of 6 11. Brief Description of Project/Program (including objectives and expected outcomes) In 2015 electricity demand reached 665.4 MW, and it is projected to reach 896 to 1,038 MW by 2026. -

Countries at the Crossroads 2012: Nicaragua
Countries at the Crossroads COUNTRIES AT THE CROSSROADS 2012: NICARAGUA INTRODUCTION Nicaragua’s November 2011 elections marked a major step forward in President Daniel Ortega’s consolidation of power, and a served as a stark demonstration of the authoritarian tendencies he has exhibited since his return to office in early 2007. A onetime guerrilla leader and the head of the Sandinista National Liberation Front (FSLN) during the1979–1990 Sandinista revolution, Ortega has ruled Nicaragua with increasing disrespect for the constitution, electoral integrity, and the rule of law. In order to run again, he engineered a questionable ruling from the Supreme Court of Justice (CSJ) to eviscerate a constitutional ban on successive terms for sitting presidents, as well as a limitation to two total terms of office.1 Ortega then secured 62 percent of the popular vote, although irregularities were widespread enough to cast doubt on the size of his victory margin. The disputed election also gave his party, the FSLN, 63 of 92 seats in the National Assembly, a majority large enough to pass ordinary legislation, change the constitution, and even call a constitutional assembly. Armed with this supermajority, Ortega is now in a position to govern in the temperamental manner of his ideological brethren in the Bolivarian Alliance for the Americas (ALBA) headed by Venezuelan president Hugo Chávez, who has provided him an economic lifeline since 2007. During his first term as president (1984–1990), Ortega presided over the drafting of a constitution in 1987 that reflected the quasi-socialist character of the revolution, which was marked by wealth redistribution and widespread confiscations of private property. -

A Cross-Sectional Epidemiological Study of Domestic Animals Related to Human
1 A cross-sectional epidemiological study of domestic animals related to human 2 leptospirosis cases in Nicaragua 3 Authors 4 Byron J. Floresa, Tania Pérez-Sánchezb, Héctor Fuertesb, Jessica Sheleby-Eliasa, José 5 Luis Múzquizb, William Jiróna, Christianne Duttmanna, Nabil Halaihelb. 6 7 a Department of Animal Health, School of Veterinary Medicine, Universidad Nacional 8 Autónoma de Nicaragua-León, Carretera a la Ceiba 1 Km al Este, León, Nicaragua. 9 b Department of Animal Pathology, Faculty of Veterinary Sciences. Universidad de 10 Zaragoza, Miguel Servet 177, 50013, Zaragoza, Spain. 11 12 Please address any correspondence to Dr. Byron José Flores Somarriba at 13 [email protected] 14 15 16 17 18 Abbreviations 19 Domestic animals (DA), Ministry of Health (MINSA, from its Spanish acronym), Pan 20 American Health Organization (PAHO), Microscopic Agglutination Test (MAT), 21 confidence intervals (CI) and Chi square test (X2). 22 1 23 ABSTRACT 24 Leptospirosis is one of the most extended zoonosis worldwide and humans become 25 infected most commonly through contact with the urine of carrier animals, either 26 directly or via contaminated water or soil. The aim in this study was to analyse the 27 epidemiological behaviour of Leptospira spp., from domestic animals around the sites 28 of human leptospirosis cases in Nicaragua, from 2007 through 2013. We report the 29 results of a cross-sectional epidemiological study with a non-probability sampling of 30 blood (n=3050) and urine (n=299) from Domestic Animals (DA) around the sites of 31 human leptospirosis cases in Nicaragua. We analysed data obtained through 32 Microscopic Agglutination Test (MAT), in-vitro culture, real time PCR and sequencing 33 of lfb1 locus. -

Nicaragua's Failed Revolution
NICARAGUA’S FAILED REVOLUTION THE INDIGENOUS STRUGGLE FOR SANEAMIENTO NICARAGUA’S FAILED REVOLUTION THE INDIGENOUS STRUGGLE FOR SANEAMIENTO Acknowledgements This report is dedicated to the courageous struggle of the Indigenous and Afro-descendent communities for Saneamiento in Nicaragua. The report was authored by Anuradha Mittal with contribution from Josh Mayer, research assistance by Elizabeth Fraser, Elena Teare, Naomi Maisel, Andy Currier, and Janhavi Mittal, and editorial assistance from Frederic Mousseau. Forests of the World provided the map on Current Land Use, Indigenous/Afro-descendent Territories and Protected Areas. Field research was conducted by Anuradha Mittal, with assistance from several individuals, who are not named so their long- term commitment to this work is not threatened. We are deeply grateful for their support. Names and details that could reveal the identity of those interviewed have been withheld to ensure their safety. The fear of invasions, killings, and kidnappings, that is silencing the Indigenous communities in Nicaragua, makes this report all the more urgent. We are deeply grateful to the many individual and foundation donors who make our work possible. Thank you. Views and conclusions expressed in this publication are those of the Oakland Institute alone and do not reflect opinions of the individuals and organizations that have sponsored and supported the work. Design: Amymade Graphic Design, amymade.com Cover Photo: Miskitu villagers on the banks of Rio Coco. November 2018 All photos © The Oakland Institute Publisher: The Oakland Institute is an independent policy think tank bringing fresh ideas and bold action to the most pressing social, economic, and environmental issues. This work is licensed under the Creative Commons Attribution 4.0 International License (CC BY-NC 4.0). -

Impact Evaluation of Component 1 of the Sustainability Project of the Rural Water and Sanitation Sector (Prosasr) Results 2015-2019
IMPACT EVALUATION OF COMPONENT 1 OF THE SUSTAINABILITY PROJECT OF THE RURAL WATER AND SANITATION SECTOR (PROSASR) RESULTS 2015-2019 Index SUMMARY � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 3 ACKNOWLEDGEMENTS � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 5 INTRODUCTION � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 7 THE STATE OF NICARAGUA’S RURAL WATER AND SANITATION SECTOR � � � � � � � � � � 13 CURRENT ACCESS TO WATER AND SANITATION, WATER DISTRIBUTION SYSTEMS, AND SAFELY MANAGED WATER � � � � � � � � � � � 14 ACCESS TO WATER AND SANITATION AS MEASURED BY SIASAR � � � � � � � � � � � � 15 WATER DISTRIBUTION SYSTEMS IN RURAL NICARAGUA—FACTS AND FIGURES � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 17 ACCESS TO WATER ACCORDING TO THE SUSTAINABLE DEVELOPMENT GOALS � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 21 THE SECTOR’S INSTITUTIONAL STRUCTURE � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 22 HOW AVAR TRAINING AND ARAS SUPPORT IMPROVEMENT OF INSTITUTIONAL CAPABILITIES � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 25 THE PROJECT GOAL: HIGHER SIASAR-BASED SCORES � � � � � � � � � � � � � � � � � � � � � 26 TRAINING THROUGH AVAR WORKSHOPS � � � � � � � � � � � � � � � � � � � � � � � � � � � -

International Health Regulations / Règlement
INTERNATIONAL HEALTH REGULATIONS / RÈGLEMENT SANITAIRE INTERNATIONAL Notifications of diseases received from 8 to 14 June 2001 / Notifications de maladies reçues du 8 au 14 juin 2001 Cases / Deaths Yellow fever / Fièvre jaune Cas / Décès Americas / Amériques Peru/Pérou 7.VI 82 Newly infected areas from 8 to 14 June 2001 / Zones nouvellement infectées entre le 8 et le 14 juin 2001 For criteria used in compiling this list, see No. 23, 2001, p. 180. X/ Les critères appliqués pour la compilation de cette liste sont publiés dans le No 23, 2001, p. 180.X Yellow fever / Fièvre jaune Americas / Amériques Peru/Pérou Loreto Department Iquitos District Puinahua District San Pablo District Areas removed from the infected area list between 8 and 14 June 2001 / Zones supprimées de la liste des zones infectées entre le 8 et le 14 juin 2001 For criteria used in compiling this list, see No. 23, 2001, p. 180. X/ Les critères appliqués pour la compilation de cette liste sont publiés dans le No 23, 2001, p. 180.X Cholera / Choléra Americas / Amériques Belize French Guiana / Guyane française Panama All infected areas / Toutes les zones infectées All infected areas / Toutes les zones infectées All infected areas / Toutes les zones infectées Bolivia / Bolivie Guyana / Guyane Suriname All infected areas / Toutes les zones infectées All infected areas / Toutes les zones infectées All infected areas / Toutes les zones infectées Colombia / Colombie Honduras Venezuela All infected areas / Toutes les zones infectées All infected areas / Toutes les zones infectées -

Hurricanes Eta and Iota
Central America Hurricanes Eta and Iota Humanitarian Situation Report No.6 © UNICEF/Nicaragua/2020/Espinoza Delgado Reporting Period 10 – 21 December 2021 Highlights Situation in Numbers • The situation remains dire for thousands of children and families affected by the passage of Eta and Iota hurricanes in Central America. Nearly 9.3 9.3 million million people have been affected (including 3.5 million children) and People affected by Eta and Iota thousands remain in shelters, facing heightened protection and public 3.5 million health risks. Children affected by Eta and Iota • Ensuring shelters are safe spaces for children and families; rehabilitation of schools – including those that have been used as shelters; access to healthy foods; rehabilitation of water systems and providing lifesaving 3.4 million water, sanitation and hygiene (WASH) supplies and services; and People in need (due to Eta) mainstreaming actions for prevention of sexual exploitation and abuse 1.3 million (PSEA); remain among the most pressing needs of the affected families. • UNICEF Country Offices (COs) continue increasing their field presence Children in need (due to Eta) and scaling up response. Progress in the response over the reporting period include: 646,943 • In Honduras: Over 4,600 people in shelters benefit daily from access to People to be reached* safe water, hygiene supplies and services. More than 30,000 more people, including those who have returned to their communities, are 327,605 starting to receive UNICEF-supported WASH services. In shelters, Children to be reached* UNICEF and partners are reaching more than 29,000 people with mental health and psychosocial support, and nearly 11,000 women, girls and boys have benefited from gender-based violence prevention interventions. -

Nicaragua: Floods Update N° 1 16 November 2010
DREF operation n° MDRNI003 GLIDE n° FL-2010-000106-NIC Nicaragua: Floods Update n° 1 16 November 2010 The International Federation of Red Cross and Red Crescent (IFRC) Disaster Relief Emergency Fund (DREF) is a source of un-earmarked money created by the Federation in 1985 to ensure that immediate financial support is available for Red Cross and Red Crescent emergency response. The DREF is a vital part of the International Federation’s disaster response system and increases the ability of National Societies to respond to disasters. Period covered by this update: 17 September to 27 October 2010. Summary: 242,122 Swiss francs were allocated from the IFRC’s Disaster Relief Emergency Fund (DREF) on 17 September 2010 to support the Nicaraguan Red Cross (NRC) in delivering assistance to some 1,000 families (5,000 beneficiaries). After Tropical Storm Agatha struck Nicaragua, the country was affected by continuous rains, increasing the impact on the population. More than 71,000 people are affected, of whom 9,904 are living in collective centres. As of the reporting period, the Nicaraguan Red Cross has reached 621 families with The community of Las Avellanas, Managua department, Tipitapa relief items and more than 500 with municipality received relief items on 1 October 2010. Photo source: psychosocial support. Nicaraguan Red Cross. This DREF update reflects the addition of the León, Chinandega and Jinotega departments as target areas for relief distributions. Some communities within the initial four target departments were restricted for access only by the military forces; therefore, the National Society is targeting affected households in other departments. -

Nicaragua: Joint Recontra-Recompa Forces Launch Protest Actions Deborah Tyroler
University of New Mexico UNM Digital Repository NotiCen Latin America Digital Beat (LADB) 5-1-1992 Nicaragua: Joint Recontra-recompa Forces Launch Protest Actions Deborah Tyroler Follow this and additional works at: https://digitalrepository.unm.edu/noticen Recommended Citation Tyroler, Deborah. "Nicaragua: Joint Recontra-recompa Forces Launch Protest Actions." (1992). https://digitalrepository.unm.edu/ noticen/7372 This Article is brought to you for free and open access by the Latin America Digital Beat (LADB) at UNM Digital Repository. It has been accepted for inclusion in NotiCen by an authorized administrator of UNM Digital Repository. For more information, please contact [email protected]. LADB Article Id: 061490 ISSN: 1089-1560 Nicaragua: Joint Recontra-recompa Forces Launch Protest Actions by Deborah Tyroler Category/Department: General Published: Friday, May 1, 1992 April 20: Reports from local media sources indicated that a joint group of recontras and recompas, known as "revueltos," occupied the towns of La Concordia and San Sebastian de Yali, Jinotega department. April 21: Sandinista Popular Army (EPS) spokespersons reported that 20 recontras ambushed and killed five peasants in La Patriota, 240 km. east of Managua. April 22: Customs inspector David Rivera told reporters that revueltos forced personnel at the El Espino border crossing to leave their posts. El Espino, Madriz department, is Nicaragua's principal overland crossing with Honduras. Rivera said the blockade prevented passage of about 40 trucks carrying cargo from Guatemala, Honduras and El Salvador, en route to Costa Rica and Panama. The recompa and recontra high commands issued a communique asserting that transportation blockades in Jinotega, Esteli and Matagalpa departments were a response to the government's failure to follow through on commitments made to demobilized combatants.