The Role of Molecular Scaffolds at the Active Zone in Synaptic Vesicle Distribution and Release Probability
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pharmacological Mechanisms Underlying the Hepatoprotective Effects of Ecliptae Herba on Hepatocellular Carcinoma
Hindawi Evidence-Based Complementary and Alternative Medicine Volume 2021, Article ID 5591402, 17 pages https://doi.org/10.1155/2021/5591402 Research Article Pharmacological Mechanisms Underlying the Hepatoprotective Effects of Ecliptae herba on Hepatocellular Carcinoma Botao Pan ,1 Wenxiu Pan ,2 Zheng Lu ,3 and Chenglai Xia 1,4 1Affiliated Foshan Maternity & Child Healthcare Hospital, Southern Medical University, Foshan 528000, China 2Department of Laboratory, Fifth People’s Hospital of Foshan, Foshan 528000, China 3Wuzhou Maternal and Child Health-Care Hospital, Wuzhou 543000, China 4School of Pharmaceutical Sciences, Southern Medical University, Guangzhou 510515, China Correspondence should be addressed to Zheng Lu; [email protected] and Chenglai Xia; [email protected] Received 30 January 2021; Revised 31 May 2021; Accepted 3 July 2021; Published 16 July 2021 Academic Editor: Hongcai Shang Copyright © 2021 Botao Pan et al. -is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Background. -e number of hepatocellular carcinoma (HCC) cases worldwide has increased significantly. As a traditional Chinese medicine (TCM) with a long history, Ecliptae herba (EH) has been widely used in HCC patients in China, but its hepatoprotective mechanism is still unclear. Methods. In this study, we applied a network pharmacology-based strategy and experimental ver- ification to systematically unravel the underlying mechanisms of EH against HCC. First, six active ingredients of EH were screened from the Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform (TCMSP) by the ADME method. Subsequently, 52 potential targets of 6 active ingredients acting on HCC were screened from various databases, including TCMSP, DGIdb, SwissTargetPrediction, CTD, and GeneCards. -

Inaugural Dissertation
Molecular Mechanisms that Regulate Neurotransmission and Establish Presynaptic Homeostasis at the Drosophila melanogaster Neuromuscular Junction Inaugural Dissertation to obtain the academic degree Doctor rerum naturalium (Dr. rer. nat.) submitted to the Department of Biology, Chemistry and Pharmacy of Freie Universität Berlin by Anthony William McCarthy from Dublin, Ireland February 2020 The experimental part of this thesis was conducted from September 2015 to November 2019 under the supervision of Dr. Alexander Walter at the Leibniz-Forschungsinstitut für Molekulare Pharmakologie (FMP) and at the CharitéCrossOver, Charité Campus Berlin Mitte. 1st Reviewer: Dr. Alexander Walter 2nd Reviewer: Prof. Dr. Stephan Sigrist Date of Defence: Statement of Authorship I hereby declare that I am the sole author of this thesis and that I have not used any sources or tools other than those quoted. Use of work by any other author is identified as such and appropriately referenced. Berlin, February 2020 Anthony William McCarthy Acknowledgments I would like to take the opportunity to thank all the people who have been important and made an impact on me during my time in Berlin. I would first like to thank my supervisor, Dr. Alexander Walter, for his guidance and supervision over the years. His passion for science and expert knowledge as well as approachable nature made for insightful conversation, projecting me forward with my projects. I would also like to thank Prof. Dr. Stephan Sigrist for his valuable input during thesis committee meetings and for acting as a reviewer. I am grateful to Prof. Dr. Volker Haucke, the members of his lab and all those at the FMP in Buch for the plentiful feedback I received following thesis committee meetings and progress presentations. -
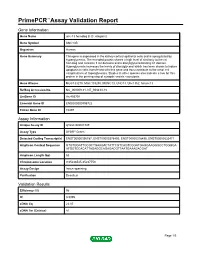
Primepcr™Assay Validation Report
PrimePCR™Assay Validation Report Gene Information Gene Name unc-13 homolog B (C. elegans) Gene Symbol UNC13B Organism Human Gene Summary This gene is expressed in the kidney cortical epithelial cells and is upregulated by hyperglycemia. The encoded protein shares a high level of similarity to the rat homolog and contains 3 C2 domains and a diacylglycerol-binding C1 domain. Hyperglycemia increases the levels of diacylglycerol which has been shown to induce apoptosis in cells transfected with this gene and thus contribute to the renal cell complications of hyperglycemia. Studies in other species also indicate a role for this protein in the priming step of synaptic vesicle exocytosis. Gene Aliases MGC133279, MGC133280, MUNC13, UNC13, Unc13h2, hmunc13 RefSeq Accession No. NC_000009.11, NT_008413.18 UniGene ID Hs.493791 Ensembl Gene ID ENSG00000198722 Entrez Gene ID 10497 Assay Information Unique Assay ID qHsaCID0007349 Assay Type SYBR® Green Detected Coding Transcript(s) ENST00000396787, ENST00000378495, ENST00000378496, ENST00000535471 Amplicon Context Sequence GTGTGGATTGCGCTGAAGACTATTCGTCAGTCGGATGAGGAAGGGCCTGGGGA ATGGTCCACATTAGAGGCAGAGACGTTAATGAAAGACGAT Amplicon Length (bp) 63 Chromosome Location 9:35236545-35237753 Assay Design Intron-spanning Purification Desalted Validation Results Efficiency (%) 96 R2 0.9995 cDNA Cq 22.07 cDNA Tm (Celsius) 81 Page 1/5 PrimePCR™Assay Validation Report gDNA Cq Specificity (%) 100 Information to assist with data interpretation is provided at the end of this report. Page 2/5 PrimePCR™Assay Validation Report -

Meta-Analysis of Pharmacogenetic Interactions in Amyotrophic Lateral Sclerosis Clinical Trials
Published Ahead of Print on October 4, 2017 as 10.1212/WNL.0000000000004606 Meta-analysis of pharmacogenetic interactions in amyotrophic lateral sclerosis clinical trials Ruben P.A. van Eijk, MD ABSTRACT Ashley R. Jones, PhD Objective: To assess whether genetic subgroups in recent amyotrophic lateral sclerosis (ALS) tri- William Sproviero, PhD als responded to treatment with lithium carbonate, but that the treatment effect was lost in a large Aleksey Shatunov, PhD cohort of nonresponders. Pamela J. Shaw, MD, PhD Methods: Individual participant data were obtained from 3 randomized trials investigating the P. Nigel Leigh, MD, PhD efficacy of lithium carbonate. We matched clinical data with data regarding the UNC13A and Carolyn A. Young, MD, C9orf72 genotype. Our primary outcome was survival at 12 months. On an exploratory basis, we PhD assessed whether the effect of lithium depended on the genotype. Christopher E. Shaw, MD, PhD Results: Clinical data were available for 518 of the 606 participants. Overall, treatment with lith- Gabriele Mora, MD ium carbonate did not improve 12-month survival (hazard ratio [HR] 1.0, 95% confidence interval – p 5 UNC13A C9orf72 Jessica Mandrioli, MD [CI] 0.7 1.4; 0.96). Both the and genotype were independent predictors – p 5 – p 5 Giuseppe Borghero, MD of survival (HR 2.4, 95% CI 1.3 4.3; 0.006 and HR 2.5, 95% CI 1.1 5.2; 0.032, UNC13A p 5 Paolo Volanti, MD respectively). The effect of lithium was different for carriers ( 0.027), but not for C9orf72 p 5 UNC13A Frank P. Diekstra, MD, carriers ( 0.22). -

Reduced Insulin Secretion Correlates with Decreased Expression of Exocytotic Genes in Pancreatic Islets from Patients with Type 2 Diabetes
Molecular and Cellular Endocrinology 364 (2012) 36–45 Contents lists available at SciVerse ScienceDirect Molecular and Cellular Endocrinology journal homepage: www.elsevier.com/locate/mce Reduced insulin secretion correlates with decreased expression of exocytotic genes in pancreatic islets from patients with type 2 diabetes Sofia A. Andersson a, Anders H. Olsson b, Jonathan L.S. Esguerra a, Emilia Heimann e, Claes Ladenvall c, Anna Edlund a, Albert Salehi d, Jalal Taneera c, Eva Degerman e, Leif Groop c, Charlotte Ling b, ⇑ Lena Eliasson a, a Islet Cell Exocytosis, Lund University Diabetes Centre, Department of Clinical Sciences Malmö, Lund University, Malmö, Sweden b Epigenetics and Diabetes, Lund University Diabetes Centre, Department of Clinical Sciences Malmö, Lund University, Malmö, Sweden c Diabetes and Endocrinology, Lund University Diabetes Centre, Department of Clinical Sciences Malmö, Lund University, Malmö, Sweden d Islet Cell Physiology, Lund University Diabetes Centre, Department of Clinical Sciences Malmö, Lund University, Malmö, Sweden e Department of Experimental Medical Sciences, Biomedical Center, Lund University, Lund, Sweden article info abstract Article history: Reduced insulin release has been linked to defect exocytosis in b-cells. However, whether expression of Received 14 December 2011 genes suggested to be involved in the exocytotic process (exocytotic genes) is altered in pancreatic islets Received in revised form 7 August 2012 from patients with type 2 diabetes (T2D), and correlate to insulin secretion, needs to be further investi- Accepted 13 August 2012 gated. Available online 23 August 2012 Analysing expression levels of 23 exocytotic genes using microarray revealed reduced expression of five genes in human T2D islets (v2 = 13.25; p < 0.001). -

Genomic and Transcriptome Analysis Revealing an Oncogenic Functional Module in Meningiomas
Neurosurg Focus 35 (6):E3, 2013 ©AANS, 2013 Genomic and transcriptome analysis revealing an oncogenic functional module in meningiomas XIAO CHANG, PH.D.,1 LINGLING SHI, PH.D.,2 FAN GAO, PH.D.,1 JONATHAN RUssIN, M.D.,3 LIYUN ZENG, PH.D.,1 SHUHAN HE, B.S.,3 THOMAS C. CHEN, M.D.,3 STEVEN L. GIANNOTTA, M.D.,3 DANIEL J. WEISENBERGER, PH.D.,4 GAbrIEL ZADA, M.D.,3 KAI WANG, PH.D.,1,5,6 AND WIllIAM J. MAck, M.D.1,3 1Zilkha Neurogenetic Institute, Keck School of Medicine, University of Southern California, Los Angeles, California; 2GHM Institute of CNS Regeneration, Jinan University, Guangzhou, China; 3Department of Neurosurgery, Keck School of Medicine, University of Southern California, Los Angeles, California; 4USC Epigenome Center, Keck School of Medicine, University of Southern California, Los Angeles, California; 5Department of Psychiatry, Keck School of Medicine, University of Southern California, Los Angeles, California; and 6Division of Bioinformatics, Department of Preventive Medicine, Keck School of Medicine, University of Southern California, Los Angeles, California Object. Meningiomas are among the most common primary adult brain tumors. Although typically benign, roughly 2%–5% display malignant pathological features. The key molecular pathways involved in malignant trans- formation remain to be determined. Methods. Illumina expression microarrays were used to assess gene expression levels, and Illumina single- nucleotide polymorphism arrays were used to identify copy number variants in benign, atypical, and malignant me- ningiomas (19 tumors, including 4 malignant ones). The authors also reanalyzed 2 expression data sets generated on Affymetrix microarrays (n = 68, including 6 malignant ones; n = 56, including 3 malignant ones). -

Whole-Genome Sequencing Reveals Genetic Variation in the Asian House Rat
INVESTIGATION Whole-Genome Sequencing Reveals Genetic Variation in the Asian House Rat Huajing Teng,*,†,‡ Yaohua Zhang,* Chengmin Shi,*,§ Fengbiao Mao,‡ Lingling Hou,‡ Hongling Guo,*,† Zhongsheng Sun,‡ and Jianxu Zhang*,1 *The State Key Laboratory of Integrated Management of Pest Insects and Rodents, Institute of Zoology, Chinese Academy of Sciences, 100101 Beijing, China, †University of Chinese Academy of Sciences, 100049 Beijing, China, § ‡ Beijing Institutes of Life Science, Chinese Academy of Sciences, 100101 Beijing, China, and Beijing Institute of Genomics, Chinese Academy of Sciences, 100101 Beijing, China ABSTRACT Whole-genome sequencing of wild-derived rat species can provide novel genomic resources, KEYWORDS which may help decipher the genetics underlying complex phenotypes. As a notorious pest, reservoir of human genetic pathogens, and colonizer, the Asian house rat, Rattus tanezumi, is successfully adapted to its habitat. However, landscape little is known regarding genetic variation in this species. In this study, we identified over 41,000,000 single- next-generation nucleotide polymorphisms, plus insertions and deletions, through whole-genome sequencing and bioinfor- sequencing matics analyses. Moreover, we identified over 12,000 structural variants, including 143 chromosomal inversions. Rattus tanezumi Further functional analyses revealed several fixed nonsense mutations associated with infection and immunity- single-nucleotide related adaptations, and a number of fixed missense mutations that may be related to anticoagulant resistance. polymorphisms A genome-wide scan for loci under selection identified various genes related to neural activity. Our whole- structural genome sequencing data provide a genomic resource for future genetic studies of the Asian house rat species variations and have the potential to facilitate understanding of the molecular adaptations of rats to their ecological niches. -

The Association of UNC13B Gene Polymorphisms and Diabetic
CLINICAL RESEARCH e-ISSN 1643-3750 © Med Sci Monit, 2019; 25: 8527-8533 DOI: 10.12659/MSM.919930 Received: 2019.09.05 Accepted: 2019.10.21 The Association of UNC13B Gene Polymorphisms Published: 2019.11.12 and Diabetic Kidney Disease in a Chinese Han Population Authors’ Contribution: AEG 1 Ya Wang 1 Department of Endocrinology, Jingzhou First People’s Hospital, The First Affiliated Study Design A AC 2 Jie Tan Hospital of Yangtze University, Jingzhou, Hubei, P.R. China Data Collection B 2 Department of Hematology, Jingzhou First People’s Hospital, The First Affiliated Statistical Analysis C BF 1 Dan Liu Hospital of Yangtze University, Jingzhou, Hubei, P.R .China Data Interpretation D BD 3 Yameng Yang 3 Department of Rheumatism and Immunology, Jingzhou First People’s Hospital, Manuscript Preparation E CG 1 Hongyan Wu The First Affiliated Hospital of Yangtze University, Jingzhou, Hubei, P.R. China Literature Search F Funds Collection G Corresponding Authors: Jie Tan, e-mail: [email protected], Hongyan Wu, e-mail: [email protected] Source of support: This study was supported by the Science and Technology Planning Project of Jingzhou Science and Technology Bureau, China (Grant No. 2018073) Background: Polymorphisms in the UNC13B gene are associated with diabetic kidney disease (DKD) in the European pop- ulation. Asian populations are more likely to suffer from complications of type 2 diabetes mellitus (T2DM), including diabetic kidney disease (DKD). This case-control study aimed to investigate the association between UNC13B gene polymorphisms and DKD in a Chinese Han population. Material/Methods: Five single nucleotide polymorphism (SNP) loci (rs13293564, rs17360668, rs10114937, rs661712, and rs2281999) were genotyped in the UNC13B gene in 600 Chinese Han subjects. -

High-Throughput Drug Screens for Amyotrophic Lateral Sclerosis Drug Discovery
This is a repository copy of High-throughput drug screens for amyotrophic lateral sclerosis drug discovery. White Rose Research Online URL for this paper: http://eprints.whiterose.ac.uk/137336/ Version: Accepted Version Article: McGown, A. orcid.org/0000-0002-3213-1895 and Stopford, M.J. orcid.org/0000-0002-1298-4102 (2018) High-throughput drug screens for amyotrophic lateral sclerosis drug discovery. Expert Opinion on Drug Discovery. ISSN 1746-0441 https://doi.org/10.1080/17460441.2018.1533953 This is an Accepted Manuscript of an article published by Taylor & Francis in Expert Opinion on Drug Discovery on 13/10/18, available online: http://www.tandfonline.com/10.1080/17460441.2018.1533953 Reuse Items deposited in White Rose Research Online are protected by copyright, with all rights reserved unless indicated otherwise. They may be downloaded and/or printed for private study, or other acts as permitted by national copyright laws. The publisher or other rights holders may allow further reproduction and re-use of the full text version. This is indicated by the licence information on the White Rose Research Online record for the item. Takedown If you consider content in White Rose Research Online to be in breach of UK law, please notify us by emailing [email protected] including the URL of the record and the reason for the withdrawal request. [email protected] https://eprints.whiterose.ac.uk/ Title: High-throughput drug screens for amyotrophic lateral sclerosis drug discovery Abstract: Amyotrophic lateral sclerosis (ALS) is a rapid adult onset neurodegenerative disorder characterised by the progressive loss of upper and lower motor neurons. -

Munc13-1/2 (T-20): Sc-13630
SAN TA C RUZ BI OTEC HNOL OG Y, INC . Munc13-1/2 (T-20): sc-13630 BACKGROUND PRODUCT Munc13 proteins (Munc13-1, Munc13-2, and Munc13-3) make up a family of Each vial contains 200 µg IgG in 1.0 ml of PBS with < 0.1% sodium azide highly homologous synaptic molecules that bind Syntaxin, an essential medi - and 0.1% gelatin. ator of neurotransmitter release. Munc13 proteins contain phorbol ester bind - Blocking peptide available for competition studies, sc-13630 P, (100 µg ing C1- and C2-domains, which are regulatory domains for Ca 2+ , phospho - pep tide in 0.5 ml PBS containing < 0.1% sodium azide and 0.2% BSA). lipids and diacylglycerol. Munc13 proteins are primarily expressed by neu rons, except for a ubiquitously expressed Munc13-2 splice variant. Munc13-1 is APPLICATIONS expressed by most neurons; it interacts with the N-terminal of Doc2 α, which is concentrated on the synaptic vesicle. Munc13-1 also interacts directly with Munc13-1/2 (T-20) is recommended for detection of Munc13-1 and Munc13- 2 msec7-1 to co-localize the two proteins at the active zone, a presynaptic, of mouse, rat and human origin by Western Blotting (starting dilution 1:200, subcellular compartment with extremely high membrane turnover. Munc13-1 dilution range 1:100-1:1000), immunofluorescence (starting dilution 1:50, is essential for synaptic vesicle maturation and plays a role in the central dilution range 1:50-1:500) and solid phase ELISA (starting dilution 1:30, priming function in synaptic vesicle exocytosis from glutamatergic synapses. -

Starting a Molecular Systems View of Endocytosis
ANRV356-CB24-20 ARI 3 September 2008 19:11 ANNUAL Protein Kinases: Starting REVIEWS Further Click here for quick links to Annual Reviews content online, a Molecular Systems View including: • Other articles in this volume of Endocytosis • Top cited articles • Top downloaded articles • Our comprehensive search Prisca Liberali, Pauli Ram¨ o,¨ and Lucas Pelkmans Institute of Molecular Systems Biology, ETH Zurich, CH-8093 Zurich, Switzerland; email: [email protected] Annu. Rev. Cell Dev. Biol. 2008. 24:501–23 Key Words First published online as a Review in Advance on membrane trafficking, phosphorylation, signal transduction, July 3, 2008 complexity, nonlinear systems, genetical physics The Annual Review of Cell and Developmental Biology is online at cellbio.annualreviews.org Abstract This article’s doi: The field of endocytosis is in strong need of formal biophysical model- 10.1146/annurev.cellbio.041008.145637 ing and mathematical analysis. At the same time, endocytosis must be Copyright c 2008 by Annual Reviews. much better integrated into cellular physiology to understand the for- by Universitat Zurich- Hauptbibliothek Irchel on 04/05/13. For personal use only. All rights reserved mer’s complex behavior in such a wide range of phenotypic variations. Annu. Rev. Cell Dev. Biol. 2008.24:501-523. Downloaded from www.annualreviews.org 1081-0706/08/1110-0501$20.00 Furthermore, the concept that endocytosis provides the space-time for signal transduction can now be experimentally addressed. In this review, we discuss these principles and argue for a systematic and top-down ap- proach to study the endocytic membrane system. We provide a summary of published observations on protein kinases regulating endocytic ma- chinery components and discuss global unbiased approaches to further map out kinase regulatory networks. -

Snareopathies: Diversity in Mechanisms and Symptoms
VU Research Portal SNAREopathies Verhage, Matthijs; Sørensen, Jakob B. published in Neuron 2020 DOI (link to publisher) 10.1016/j.neuron.2020.05.036 document version Publisher's PDF, also known as Version of record document license Article 25fa Dutch Copyright Act Link to publication in VU Research Portal citation for published version (APA) Verhage, M., & Sørensen, J. B. (2020). SNAREopathies: Diversity in Mechanisms and Symptoms. Neuron, 107(1), 22-37. https://doi.org/10.1016/j.neuron.2020.05.036 General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. • Users may download and print one copy of any publication from the public portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the public portal ? Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. E-mail address: [email protected] Download date: 29. Sep. 2021 ll Review SNAREopathies: Diversity in Mechanisms and Symptoms Matthijs Verhage1,2,* and Jakob B. Sørensen3,* 1Department of Functional Genomics, Vrije Universiteit (VU) Amsterdam, De Boelelaan 1085, Amsterdam 1081 HV, the Netherlands 2Department of Clinical Genetics, UMC Amsterdam, De Boelelaan 1085, Amsterdam 1081 HV, the Netherlands 3Department of Neuroscience, University of Copenhagen, 2200 Copenhagen N, Denmark *Correspondence: [email protected] (M.V.), [email protected] (J.B.S.) https://doi.org/10.1016/j.neuron.2020.05.036 Neuronal SNAREs and their key regulators together drive synaptic vesicle exocytosis and synaptic transmis- sion as a single integrated membrane fusion machine.