FINAL REPORT INCA PROJECT NO. 223 the BIOLOGICAL IMPORTANCE of COPPER a Literature Review June, 1989
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

G2 Phase Cell Cycle Regulation by E2F4 Following Genotoxic Stress
G2 Phase Cell Cycle Regulation by E2F4 Following Genotoxic Stress by MEREDITH ELLEN CROSBY Submitted in partial fulfillment of the requirements for the Degree of Doctor of Philosophy Thesis Advisor: Dr. Alex Almasan Department of Environmental Health Sciences CASE WESTERN RESERVE UNIVERSITY May, 2006 CASE WESTERN RESERVE UNIVERSITY SCHOOL OF GRADUATE STUDIES We hereby approve the dissertation of ______________________________________________________ candidate for the Ph.D. degree *. (signed)_______________________________________________ (chair of the committee) ________________________________________________ ________________________________________________ ________________________________________________ ________________________________________________ ________________________________________________ (date) _______________________ *We also certify that written approval has been obtained for any proprietary material contained therein. TABLE OF CONTENTS TABLE OF CONTENTS………………………………………………………………….1 LIST OF FIGURES……………………………………………………………………….5 LIST OF TABLES………………………………………………………………………...7 ACKNOWLEDGEMENTS……………………………………………………………….8 LIST OF ABBREVIATIONS……………………………………………………………10 ABSTRACT……………………………………………………………………………...15 CHAPTER 1. INTRODUCTION 1.1. CELL CYCLE REGULATION: HISTORICAL OVERVIEW………………...17 1.2. THE E2F FAMILY OF TRANSCRIPTION FACTORS……………………….22 1.3. E2F AND CELL CYCLE CONTROL 1.3.1. G0/G1 Phase Transition………………………………………………….28 1.3.2. S Phase…………………………………………………………………...28 1.3.3. G2/M Phase Transition…………………………………………………..30 1.4. -

Individual Index
INDIVIDUAL INDEX Bl Page Page A Cooremans-Cruz, Eugenio Juan 3825 Cortes, Eugenia 2474 Abbott, Charles P 3807 Crowley, Daniel 3820 Abrams, Samuel S 3825 Cruz, Marinelle Khristy 3836 Abshire, Arthur J 3812 Cuervo, Ester 3825 Ah Kwang Chang 3827 Cuffy, Jackson Ormiston Edwards and Ah Sing Ying 3826 Merle Cleopatra Edwards 38J54 Ah Young Cho Kwak 3803 Alcala-Salcedo, Apolinario 3825 D Alvarado, Irma Victoria Bolarte 3853 Araujo,Lilia 3828 D'Souza, Eustace John 3824 Araujo, Maria Freitas 3825 Datronics Engineers, Inc 3857 Arellano, Santos Marquez 3830 Daves, Gary 3862 Armstrong, Anne L 235 Davies, Mrs. Olive M.V.T., Samira Arriola, Rodolfo N 3851 D.K., Ola-Tomi K., Ola-Yinka K., Asencio-Placencio, Pedro 3825 Ilesha E.K., and Baba-Tunji K 3803 Atherley, Juana Todd 3829 Davis, Jefferson F 1304 Austin, John P 234 Day, Craig 3824 DeBravo, Cenovia Mesa 3825 B DeLuna-Segovia, Jose 3825 Dibben, James William 3823 Bader, Louis William 3825 Din, Ruben P 3831 Baglieri, George 3825 Do Sook Park 3841 Bakierowska, Kazimiera 3827 Dom Min Lee 3826 Bally, George Ally 3827 Dowd, Kazuko Nishioka 3820 Barengo, Bernardino 2485 Barrera-Cabrera, Jesus 3825 Batavia Turf Farms, Inc 3818 • ^- • E Benney, Donna Marainne 3836 Eaker, Lt. Gen. IraC 1060 Berger, Harry 3825 Eha,Elmar 3825 Binabise, Juanita 3840 Ehard,Karin 3858 Birely, Victor M 1643 Elder, William J 3814 Blakeley, Hildegard G 3845 Emde, Noel Abueg 3837 Blanquicett, Carmen Cecilia 3859 Encomienda, Rogelio M 3853 Bonderenko, Edward J 2485 Espenueva, Edmundo Alfredo Boone, Master Sgt. William E 3813 Oreiro -

Global Invasion History of the Agricultural Pest Butterfly Pieris Rapae Revealed with Genomics and Citizen Science
Global invasion history of the agricultural pest butterfly Pieris rapae revealed with genomics and citizen science Sean F. Ryana,b,1, Eric Lombaertc, Anne Espesetd, Roger Vilae, Gerard Talaverae,f,g, Vlad Dinca˘ h, Meredith M. Doellmani, Mark A. Renshawj, Matthew W. Engi, Emily A. Hornettk,l, Yiyuan Lii, Michael E. Pfrenderi,m, and DeWayne Shoemakera,1 aDepartment of Entomology and Plant Pathology, University of Tennessee, Knoxville, TN 37996; bEcological and Biological Sciences Practice, Exponent, Inc., Menlo Park, CA 94025; cInstitut Sophia Agrobiotech, Centre de Recherches de Sophia-Antipolis, Université Côte d’Azur, Centre National de la Recherche Scientifique, Institut National de la Recherche Agronomique, 06 903 Sophia Antipolis, France; dDepartment of Biology, University of Nevada, Reno, NV 89557; eDepartment of Animal Biodiversity and Evolution, Institut de Biologia Evolutiva, Consejo Superior de Investigaciones Científicas and Universitat Pompeu Fabra, Barcelona, 08003 Spain; fDepartment of Organismic and Evolutionary Biology, Harvard University, Cambridge, MA 02138; gMuseum of Comparative Zoology, Harvard University, Cambridge, MA 02138; hDepartment of Ecology and Genetics, University of Oulu, Oulu, 90014 Finland; iDepartment of Biological Sciences, University of Notre Dame, Notre Dame, IN 46556; jShrimp Department, Oceanic Institute, Hawai’i Pacific University, Waimanalo, HI 96795; kDepartment of Evolution, Ecology and Behaviour, University of Liverpool, L69 7ZB Liverpool, United Kingdom; lDepartment of Vector Biology, Liverpool -

Contribution to the Lepidopterans of Visakhapatnam Region, Andhra Pradesh, India
ANALYSIS Vol. 21, Issue 68, 2020 ANALYSIS ARTICLE ISSN 2319–5746 EISSN 2319–5754 Species Contribution to the Lepidopterans of Visakhapatnam Region, Andhra Pradesh, India Solomon Raju AJ1, Venkata Ramana K2 1Department of Environmental Sciences, Andhra University, Visakhapatnam 530 003, India 2Department of Botany, Andhra University, Visakhapatnam 530 003, India Corresponding author: A.J. Solomon Raju, Department of Environmental Sciences, Andhra University, Visakhapatnam 530 003, India Mobile: 91-9866256682, email: [email protected] Article History Received: 28 June 2020 Accepted: 02 August 2020 Published: August 2020 Citation Solomon Raju AJ, Venkata Ramana K. Contribution to the Lepidopterans of Visakhapatnam Region, Andhra Pradesh, India. Species, 2020, 21(68), 275-280 Publication License This work is licensed under a Creative Commons Attribution 4.0 International License. General Note Article is recommended to print as color digital version in recycled paper. ABSTRACT The butterflies Byblia ilithyia (Nymphalidae), Pieris canidia (Pieridae) and Azanus jesous (Lycaenidae) and the day-flying moth, Nyctemera adversata (Erebidae) are oligophagous. Previously, only B. ilithyia has been reported to be occurring in this region while the other three species are being reported for the first time from this region. The larval host plants include Jatropha gossypiifolia and Tragia involucrata for B. ilithyia, Brassica oleracea var. oleracea and B. oleracea var. botrytis for P. canidia, and Acacia auriculiformis for Azanus jesous. The nectar plants include Tragia involucrata, Euphorbia hirta and Jatropha gossypiifolia for B. ilithyia, Premna latifolia and Cleome viscosa for P. canidia, Lagascea mollis, Tridax procumbens and Digera muricata for A. jesous and Bidens pilosa for 275 N. adversata. The study recommends extensive field investigations to find out more larval plants and nectar plants for each Page lepidopteran species now reported. -
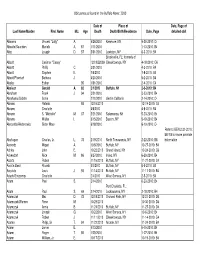
Obituaries Buffalo News 2010 by Name
Obituaries as found in the Buffalo News: 2010 Date of Place of Date, Page of Last Name/Maiden First Name M.I. Age Death Death/Birth/Residence Date, Page detailed obit Abbarno Vincent "Lolly" A. 9/26/2010 Kenmore, NY 9-30-2010: C4 Abbatte/Saunders Murielle A. 87 1/11/2010 1-13-2010: B4 Abbo Joseph D. 57 5/31/2010 Lewiston, NY 6-3-2010: B4 Brooksville, FL; formerly of Abbott Casimer "Casey" 12/19/22009 Cheektowaga, NY 4-18-2010: C6 Abbott Phillip C. 3/31/2010 4-3-2010: B4 Abbott Stephen E. 7/6/2010 7-8-2010: B4 Abbott/Pfoetsch Barbara J. 4/20/2010 5-2-2010: B4 Abeles Esther 95 1/31/2010 2-4-2010: C4 Abelson Gerald A. 82 2/1/2010 Buffalo, NY 2-3-2010: B4 Abraham Frank J. 94 3/21/2010 3-23-2010: B4 Abrahams/Gichtin Sonia 2/10/2010 died in California 2-14-2010: C4 Abramo Rafeala 93 12/16/2010 12-19-2010: C4 Abrams Charlotte 4/6/2010 4-8-2010: B4 Abrams S. "Michelle" M. 37 5/21/2010 Salamanca, NY 5-23-2010: B4 Abrams Walter I. 5/15/2010 Basom, NY 5-19-2010: B4 Abrosette/Aksterowicz Sister Mary 6/18/2010 6-19-2010: C4 Refer to BEN 2-21-2010: B6/7/8 for more possible Abshagen Charles, Jr. L. 73 2/19/2010 North Tonawanda, NY 2-22-2010: B8 information Acevedo Miguel A. 10/6/2010 Buffalo, NY 10-27-2010: B4 Achkar John E. -

Men Final Entries
Final Entries - Athletes List by event European Athletics Indoor Championships 2021, Torun/POL Tot. Number of countries Tot. Number of athletes Tot. Number of Men Tot. Number of Women 47 733 405 328 FINAL ENTRIES - Men 60m Senior Men Num. of countries: 33 Num. of athletes: 71 Member Federation Surname First Name DoB PB SB ARM Donigian Alexander 20/10/1993 6.64i 6.79i ART Keletela Dorian Celeste 06/02/1999 6.79i 6.85i AUT Fuchs Markus 14/11/1995 6.62i 6.69i BEL Kuba Di-Vita Gaylord 17/11/1995 6.73i 6.75i BEL Vleminckx Kobe 31/05/1998 6.65i 6.65i BLR Bliznets Dzianis 12/03/1995 6.75i 6.75i BLR Bohdan Maksim 19/03/1997 6.77i 6.77i BLR Zabalotny Yury 24/02/1997 6.72i 6.72i BUL Dimitrov Denis 10/02/1994 6.65i 6.73i BUL Jivkov Vesselin 26/01/2001 6.76i 6.80i CZE Hampl Štěpán 10/11/1999 6.70i 6.70i CZE Stromšík Zdeněk 25/11/1994 6.60i 6.68i CZE Veleba Jan 06/12/1986 6.65i 6.65i DEN Hansen Simon 30/06/1998 6.75i 6.75i DEN Kjær Emil Mader 20/12/1999 6.77i 6.77i DEN Musah Kojo 15/04/1996 6.61i 6.61i ESP López Sergio 05/07/1999 6.67i 6.74i ESP Rodríguez Daniel 26/01/1995 6.67i 6.67i ESP Sanchez Ricardo 10/08/1999 6.75i 6.75i EST Nazarov Karl Erik 17/03/1999 6.63i 6.63i FIN Illukka Riku 21/09/1999 6.73i 6.73i FIN Purola Samuel 19/05/2000 6.67i 6.67i FIN Samuelsson Samuli 23/06/1995 6.66i 6.66i FRA Fall Mouhamadou 25/02/1992 6.62i 6.62i FRA Golitin Amaury 28/01/1997 6.62i 6.62i GBR Aikines-Aryeetey Harry 29/08/1988 6.55i 6.67i GBR Bromby Oliver 30/03/1998 6.63i 6.65i GBR Robertson Andrew 17/12/1990 6.54i 6.61i GER Corucle Philipp 18/07/1997 6.62i -

Jammu and Kashmir) of India Anu Bala*, J
International Journal of Interdisciplinary and Multidisciplinary Studies (IJIMS), 2014, Vol 1, No.7, 24-34. 24 Available online at http://www.ijims.com ISSN: 2348 – 0343 Butterflies of family Pieridae reported from Jammu region (Jammu and Kashmir) of India Anu Bala*, J. S. Tara and Madhvi Gupta Department of Zoology, University of Jammu Jammu-180,006, India *Corresponding author: Anu Bala Abstract The present article incorporates detailed field observations of family Pieridae in Jammu region at different altitudes during spring, summer and autumn seasons of 2012-2013. The study revealed that 13 species of butterflies belonging to 10 genera of family Pieridae exist in the study area. Most members of Family Pieridae are white or yellow. Pieridae is a large family of butterflies with about 76 genera containing approximately 1,100 species mostly from tropical Africa and Asia. Keywords :Butterflies, India, Jammu, Pieridae. Introduction Jammu and Kashmir is the northernmost state of India. It consists of the district of Bhaderwah, Doda, Jammu, Kathua, Kishtwar, Poonch, Rajouri, Ramban, Reasi, Samba and Udhampur. Most of the area of the region is hilly and Pir Panjal range separates it from the Kashmir valley and part of the great Himalayas in the eastern districts of Doda and Kishtwar. The main river is Chenab. Jammu borders Kashmir to the north, Ladakh to the east and Himachal Pradesh and Punjab to the south. In east west, the line of control separates Jammu from the Pakistan region called POK. The climate of the region varies with altitude. The order Lepidoptera contains over 19,000 species of butterflies and 100,000 species of moths worldwide. -

Sales Report
Sales Listing Report Page 1 of 262 McHenry County 01/21/2021 08:25:40 01/01/2020 - 09/30/2020 Township: GRAFTON TWP Document Number Sale Year Sale Type Valid Sale Sale Date Dept. Study Selling Price Parcel Number Built Year Property Type Prop. Class Acres Square Ft. Lot Size Grantor Name Grantee Name Site Address 2020R0006895 2020 Warranty Deed Y 02/13/2020 Y $189,000.00 REINVEST HOMES, LLC ANGELA STROPE 160 S HEATHER DR 18-01-101-027 0 GARAGE/ NO O O 0040 .00 0 CRYSTAL LAKE, IL 600145177 160 S HEATHER DR CRYSTAL LAKE, IL 60014 - Legal Description: DOC 2020R0006895 LT 31 BLK 15 R A CEPEKS CRYSTAL VISTA 2020R0030830 2020 Partial Assessmen N 07/30/2020 N $136,500.00 DANIEL M ADAMS, AS TRUST BRETT A RIDINGS 1324 THORNWOOD LN 18-01-102-037 0 GARAGE/ NO O O 0040 .00 0 CRYSTAL LAKE, IL 600145042 TARI D RIDINGS 1324 THORNWOOD LANE CRYSTAL LAKE, IL 600140000 1324 THORNWOOD LN CRYSTAL LAKE, IL 60014 -5042 Legal Description: DOC 2020R0030830 LT 1 & PT LT 19 LYING N OF & ADJ BLK 14 R A CEPEKS CRYSTAL VISTA 2020R0010709 2020 Warranty Deed Y 03/16/2020 Y $210,000.00 PATRICK J. CARAMELA JUAN P. MIRANDA STACY C. CARAMELA 58 HOLLY DR 18-01-106-018 0 LDG SINGLE FAM 0040 .00 0 CRYSTAL LAKE, IL 600145022 58 HOLLY DR CRYSTAL LAKE, IL 60014 -5022 Legal Description: DOC 2020R0010709 LT 7 BLK 19 R A CEPEKS CRYSTAL VISTA 2020R0015717 2020 Warranty Deed Y 04/23/2020 Y $95,000.00 LISA M SULMA FANO THEOFANOUS 5011 NORTHWEST HWY 18-01-107-011 0 LDG SINGLE FAM 0040 .00 0 CRYSTAL LAKE, IL 600147330 STEVE THEOFANOUS 5011 NORTHWEST HIGHWAY CRYSTAL LAKE, IL 600147330 113 HEATHER DR CRYSTAL LAKE, IL 60014 - Legal Description: DOC 2020R0015717 LT 12 BLK 31 R A CEPEKS CRYSTAL VISTA Copyright (C) 1997-2021 DEVNET Incorporated MXSTIEG Sales Listing Report Page 2 of 262 McHenry County 01/21/2021 08:25:40 01/01/2020 - 09/30/2020 Township: GRAFTON TWP Document Number Sale Year Sale Type Valid Sale Sale Date Dept. -
![Arxiv:1810.00224V2 [Q-Bio.PE] 7 Dec 2020 Humanity Is Increasingly Influencing Global Environments [195]](https://docslib.b-cdn.net/cover/3556/arxiv-1810-00224v2-q-bio-pe-7-dec-2020-humanity-is-increasingly-in-uencing-global-environments-195-943556.webp)
Arxiv:1810.00224V2 [Q-Bio.PE] 7 Dec 2020 Humanity Is Increasingly Influencing Global Environments [195]
A Survey of Biodiversity Informatics: Concepts, Practices, and Challenges Luiz M. R. Gadelha Jr.1* Pedro C. de Siracusa1 Artur Ziviani1 Eduardo Couto Dalcin2 Helen Michelle Affe2 Marinez Ferreira de Siqueira2 Luís Alexandre Estevão da Silva2 Douglas A. Augusto3 Eduardo Krempser3 Marcia Chame3 Raquel Lopes Costa4 Pedro Milet Meirelles5 and Fabiano Thompson6 1National Laboratory for Scientific Computing, Petrópolis, Brazil 2Friedrich-Schiller-University Jena, Jena, Germany 2Rio de Janeiro Botanical Garden, Rio de Janeiro, Brazil 3Oswaldo Cruz Foundation, Rio de Janeiro, Brazil 4National Institute of Cancer, Rio de Janeiro, Brazil 5Federal University of Bahia, Salvador, Brazil 6Federal University of Rio de Janeiro, Rio de Janeiro, Brazil Abstract The unprecedented size of the human population, along with its associated economic activities, have an ever increasing impact on global environments. Across the world, countries are concerned about the growing resource consumption and the capacity of ecosystems to provide them. To effectively conserve biodiversity, it is essential to make indicators and knowledge openly available to decision-makers in ways that they can effectively use them. The development and deployment of mechanisms to produce these indicators depend on having access to trustworthy data from field surveys and automated sensors, biological collections, molec- ular data, and historic academic literature. The transformation of this raw data into synthesized information that is fit for use requires going through many refinement steps. The methodologies and techniques used to manage and analyze this data comprise an area often called biodiversity informatics (or e-Biodiversity). Bio- diversity data follows a life cycle consisting of planning, collection, certification, description, preservation, discovery, integration, and analysis. -
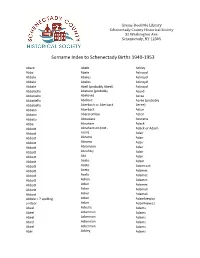
Surname Index to Schenectady Births 1940-1953
Grems-Doolittle Library Schenectady County Historical Society 32 Washington Ave. Schenectady, NY 12305 Surname Index to Schenectady Births 1940-1953 Abare Abele Ackley Abba Abele Ackroyd Abbale Abeles Ackroyd Abbale Abeles Ackroyd Abbale Abell (probably Abeel) Ackroyd Abbatiello Abelone (probably Acord Abbatiello Abelove) Acree Abbatiello Abelove Acree (probably Abbatiello Aberbach or Aberback Aeree) Abbato Aberback Acton Abbato Abercrombie Acton Abbato Aboudara Acucena Abbe Abraham Adack Abbott Abrahamson (not - Adack or Adach Abbott nson) Adair Abbott Abrams Adair Abbott Abrams Adair Abbott Abramson Adair Abbott Abrofsky Adair Abbott Abt Adair Abbott Aceto Adam Abbott Aceto Adamczak Abbott Aceto Adamec Abbott Aceto Adamec Abbott Acken Adamec Abbott Acker Adamec Abbott Acker Adamek Abbott Acker Adamek Abbzle = ? spelling Acker Adamkiewicz unclear Acker Adamkiewicz Abeel Ackerle Adams Abeel Ackerman Adams Abeel Ackerman Adams Abeel Ackerman Adams Abeel Ackerman Adams Abel Ackley Adams Grems-Doolittle Library Schenectady County Historical Society 32 Washington Ave. Schenectady, NY 12305 Surname Index to Schenectady Births 1940-1953 Adams Adamson Ahl Adams Adanti Ahles Adams Addis Ahman Adams Ademec or Adamec Ahnert Adams Adinolfi Ahren Adams Adinolfi Ahren Adams Adinolfi Ahrendtsen Adams Adinolfi Ahrendtsen Adams Adkins Ahrens Adams Adkins Ahrens Adams Adriance Ahrens Adams Adsit Aiken Adams Aeree Aiken Adams Aernecke Ailes = ? Adams Agans Ainsworth Adams Agans Aker (or Aeher = ?) Adams Aganz (Agans ?) Akers Adams Agare or Abare = ? Akerson Adams Agat Akin Adams Agat Akins Adams Agen Akins Adams Aggen Akland Adams Aggen Albanese Adams Aggen Alberding Adams Aggen Albert Adams Agnew Albert Adams Agnew Albert or Alberti Adams Agnew Alberti Adams Agostara Alberti Adams Agostara (not Agostra) Alberts Adamski Agree Albig Adamski Ahave ? = totally Albig Adamson unclear Albohm Adamson Ahern Albohm Adamson Ahl Albohm (not Albolm) Adamson Ahl Albrezzi Grems-Doolittle Library Schenectady County Historical Society 32 Washington Ave. -

Global Journal of Science Frontier Research: C Biological Science Botany & Zology
Online ISSN : 2249-4626 Print ISSN : 0975-5896 DOI : 10.17406/GJSFR DiversityofButterflies RevisitingMelaninMetabolism InfluenceofHigh-FrequencyCurrents GeneticStructureofSitophilusZeamais VOLUME20ISSUE4VERSION1.0 Global Journal of Science Frontier Research: C Biological Science Botany & Zology Global Journal of Science Frontier Research: C Biological Science Botany & Zology Volume 20 Issue 4 (Ver. 1.0) Open Association of Research Society Global Journals Inc. © Global Journal of Science (A Delaware USA Incorporation with “Good Standing”; Reg. Number: 0423089) Frontier Research. 2020 . Sponsors:Open Association of Research Society Open Scientific Standards All rights reserved. This is a special issue published in version 1.0 Publisher’s Headquarters office of “Global Journal of Science Frontier Research.” By Global Journals Inc. Global Journals ® Headquarters All articles are open access articles distributed 945th Concord Streets, under “Global Journal of Science Frontier Research” Framingham Massachusetts Pin: 01701, Reading License, which permits restricted use. United States of America Entire contents are copyright by of “Global USA Toll Free: +001-888-839-7392 Journal of Science Frontier Research” unless USA Toll Free Fax: +001-888-839-7392 otherwise noted on specific articles. No part of this publication may be reproduced Offset Typesetting or transmitted in any form or by any means, electronic or mechanical, including G lobal Journals Incorporated photocopy, recording, or any information storage and retrieval system, without written 2nd, Lansdowne, Lansdowne Rd., Croydon-Surrey, permission. Pin: CR9 2ER, United Kingdom The opinions and statements made in this book are those of the authors concerned. Packaging & Continental Dispatching Ultraculture has not verified and neither confirms nor denies any of the foregoing and no warranty or fitness is implied. -

Iveigh Ivelan Iveland Ivelane Ivelant Ivelen Ivelend Ivelent Ivelind Ivelint
Iveigh Ivones Izearde Izzach Izzox Ivelan Ivoomb Izeart Izzack Izzoyd Iveland Ivoombe Izech Izzacke Izzsard Ivelane Ivor Izeck Izzaech Izzude Ivelant Ivors Izerd Izzaeck Izzume Ivelen Ivory Izert Izzaick Izzyck Ivelend Ivvaren Izett Izzaitch Izzyke Ivelent Ivy Iziac Izzake Ivelind Ivye Izick Izzaock Ivelint Ivymey Izitch Izzaox Ivelyn Ivys Izock Izzard Ivelynd Iwadby Izod Izzarde Ivemey Iwardbey Izode Izzarn Ivemmy Iwardby Izoed Izzarne Ivemy Iwardy Izoit Izzart Iveney Iwerdby Izold Izzarte Ivens Iwoodby Izombe Izzat Iver Ix Izon Izzatt Iverach Ixton Izood Izzayck Iverack Iyon Izoode Izzayke Iveragh Iyone Izoold Izzeard Iverake Iyonne Izoomb Izzearde Iverech Izaac Izoombe Izzeart Ivereck Izaach Izord Izzech Iverick Izaack Izould Izzeck Iveritch Izaacs Izoyd Izzerd Iverock Izaacson Izrael Izzert Iverox Izaake Izraeler Izzett Ivers Izach Izraely Izzick Iverson Izack Izreeli Izzitch Ivery Izacke Izsard Izzock Iveryck Izaech Izsarde Izzod Iveryke Izaeck Izsardt Izzode Ives Izaick Izsart Izzoed Iveson Izaitch Izsarte Izzoit Ivet Izake Izseard Izzold Ivett Izaock Izsearde Izzolm Ivetts Izaox Izseart Izzomb Ivey Izard Izserd Izzombe Ivie Izarde Izsert Izzome Ivimey Izarn Izsord Izzone Ivinges Izarne Izude Izzood Ivington Izart Izyck Izzoode Ivinney Izarte Izyke Izzoold Ivins Izat Izzaac Izzoom Ivinson Izatson Izzaach Izzoomb Iviormy Izatt Izzaack Izzoombe Ivison Izayck Izzaacs Izzord Ivombe Izayke Izzaacson Izzould Ivone Izeard Izzaake Izzown Hall of Names by Swyrich © 1999 Swyrich Corporation www.swyrich.com 1-888-468-7686 557 Jackline Jacok