Typification of Arisarum Proboscideum
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

LD5655.V855 1924.M377.Pdf
A STUDY OF BACILLUS AROIDEAE 'l'OWNSEND, THE CAUSE OF A SOFT ROT OF TOMATO, AN:) B. CAROTOVORUS JONES. A thesis 811.bmitted as partial requirement for the degree of Master of Science in Botany by A.B.Massey Reprinted from PHYT0PATH0L0GY, Vol. XIV, October, 1924. A STUDY OF BACILLUS AROIDEAE, TOWNSEND, THE CAUSE OF A SOFT ROT OF TOMATO, AND B. CAROTOVORUS JONES A. B. MASSEY! \Vl'l'll Tlll:EE FIGURES IN THE TEXT INTRODUCTION In the summer of 1918, at Blacksburg, Virginia, there developed a con- siderable amount of a soft rot of tomatoes. This occurre<l in experimental plots which were designated to study the control of septoria leaf blight, and the soft rot of the fruit developed into an important factor. I~ describing these experiments Fromme (2) states: "Practically all of the unsoundness of the fruit was caused by bacterial soft rot, a disease which is exceedingly common and often very destructive in tomato fields in Vir- ginia." Isolations from diseased fruits made by S. A. Wingard (15) proved a bacterium to be the causative agent. Its growth in pure culture resembled that of the group of bacteria which causes soft rots of plants but it could not be readily as!ligned to any of the describe<l species of this group. There has been only casual mention of a bacterial soft rot of tomato in literature, and the distinguishing features of the organisms which might be responsible have not been as sharply defined as is desirable. It was decided, therefore, to un<lcrtake comparative studies of the organism in question together with some of the non-chromogenic soft rot forms. -

Leaf Variegation in Caladium Steudneriifolium (Araceae): a Case of Mimicry?
Evol Ecol (2009) 23:503–512 DOI 10.1007/s10682-008-9248-2 ORIGINAL PAPER Leaf variegation in Caladium steudneriifolium (Araceae): a case of mimicry? Ulf Soltau Æ Stefan Do¨tterl Æ Sigrid Liede-Schumann Received: 27 November 2007 / Accepted: 25 February 2008 / Published online: 6 March 2008 Ó Springer Science+Business Media B.V. 2008 Abstract The leaves of Caladium steudneriifolium (Araceae) of the understorey of a submontane rainforest in the Podocarpus National Park (South East Ecuador, 1,060 m a.s.l.) are plain green or patterned with whitish variegation. Of the 3,413 individual leaves randomly chosen and examined in April 2003, two-thirds were plain green, whereas one third were variegated (i.e., whitish due to absence of chloroplasts). Leaves of both morphs are frequently attacked by mining moth caterpillars. Our BLAST analysis based on Cytochrome-c-Oxidase-subunit-1 sequences suggests that the moth is possibly a member of the Pyraloidea or another microlepidopteran group. It was observed that the variegated leaf zones strongly resemble recent damages caused by mining larvae and therefore may mimic an attack by moth larvae. Infestation was significantly 4–12 times higher for green leaves than for variegated leaves. To test the hypothesis that variegation can be interpreted as mimicry to deter ovipositing moths, we first ruled out the possibility that variegation is a function of canopy density (i.e., that the moths might be attracted or deterred by factors unrelated to the plant). Then plain green leaves were artificially variegated and the number of mining larvae counted after 3 months. -

My Green Wet Thumb: Lagenandra
My Green Wet Thumb: Lagenandra By Derek P.S. Tustin Over the years I have found that the average aquarist will go through several different stages. I am by no means a sociologist specializing in the aquari- um hobbyist, but from my own observations I think pretty much everyone goes through some variation of the following; Initial wide-spread interest and associated errors, A focusing of interest into one or two main areas, Competence in an area of interest, Mastery of an area of interest Expansion of interest into new areas while either maintaining the old interest, or focusing entirely on the new area of interest. As an aquatic horticulturist, there are actually very few entry points, or at least entry species, into the hobby. When I started out, I had access to sev- eral excellent aquarium stores with an impressive diversity of aquatic crea- tures, but a very limited selection of aquatic plants. Now, this was back be- fore I joined the Durham Region Aquarium Society (DRAS), so I didn’t have access to mentors or their specialized stock, and it was also before there were so many excellent on-line resources. Most of my initial experience came from the limited genera of plants that were available in local stores; Echinodorus, Cryptocoryne, Anubias and some Aponogeton. (Oh, there were numerous stem plants, but for some reason, I have never been that interested in those, being much more fascinated by rooted plants, and my interest in ponds and suitable plants came much later.) Over the past decade, I have grown the majority of commonly available plants from those genera, and now also have the benefit of being exposed to other skilled hobbyists and resources offered through DRAS. -

Devonian Plant Fossils a Window Into the Past
EPPC 2018 Sponsors Academic Partners PROGRAM & ABSTRACTS ACKNOWLEDGMENTS Scientific Committee: Zhe-kun Zhou Angelica Feurdean Jenny McElwain, Chair Tao Su Walter Finsinger Fraser Mitchell Lutz Kunzmann Graciela Gil Romera Paddy Orr Lisa Boucher Lyudmila Shumilovskikh Geoffrey Clayton Elizabeth Wheeler Walter Finsinger Matthew Parkes Evelyn Kustatscher Eniko Magyari Colin Kelleher Niall W. Paterson Konstantinos Panagiotopoulos Benjamin Bomfleur Benjamin Dietre Convenors: Matthew Pound Fabienne Marret-Davies Marco Vecoli Ulrich Salzmann Havandanda Ombashi Charles Wellman Wolfram M. Kürschner Jiri Kvacek Reed Wicander Heather Pardoe Ruth Stockey Hartmut Jäger Christopher Cleal Dieter Uhl Ellen Stolle Jiri Kvacek Maria Barbacka José Bienvenido Diez Ferrer Borja Cascales-Miñana Hans Kerp Friðgeir Grímsson José B. Diez Patricia Ryberg Christa-Charlotte Hofmann Xin Wang Dimitrios Velitzelos Reinhard Zetter Charilaos Yiotis Peta Hayes Jean Nicolas Haas Joseph D. White Fraser Mitchell Benjamin Dietre Jennifer C. McElwain Jenny McElwain Marie-José Gaillard Paul Kenrick Furong Li Christine Strullu-Derrien Graphic and Website Design: Ralph Fyfe Chris Berry Peter Lang Irina Delusina Margaret E. Collinson Tiiu Koff Andrew C. Scott Linnean Society Award Selection Panel: Elena Severova Barry Lomax Wuu Kuang Soh Carla J. Harper Phillip Jardine Eamon haughey Michael Krings Daniela Festi Amanda Porter Gar Rothwell Keith Bennett Kamila Kwasniewska Cindy V. Looy William Fletcher Claire M. Belcher Alistair Seddon Conference Organization: Jonathan P. Wilson -

The Evolution of Pollinator–Plant Interaction Types in the Araceae
BRIEF COMMUNICATION doi:10.1111/evo.12318 THE EVOLUTION OF POLLINATOR–PLANT INTERACTION TYPES IN THE ARACEAE Marion Chartier,1,2 Marc Gibernau,3 and Susanne S. Renner4 1Department of Structural and Functional Botany, University of Vienna, 1030 Vienna, Austria 2E-mail: [email protected] 3Centre National de Recherche Scientifique, Ecologie des Foretsˆ de Guyane, 97379 Kourou, France 4Department of Biology, University of Munich, 80638 Munich, Germany Received August 6, 2013 Accepted November 17, 2013 Most plant–pollinator interactions are mutualistic, involving rewards provided by flowers or inflorescences to pollinators. An- tagonistic plant–pollinator interactions, in which flowers offer no rewards, are rare and concentrated in a few families including Araceae. In the latter, they involve trapping of pollinators, which are released loaded with pollen but unrewarded. To understand the evolution of such systems, we compiled data on the pollinators and types of interactions, and coded 21 characters, including interaction type, pollinator order, and 19 floral traits. A phylogenetic framework comes from a matrix of plastid and new nuclear DNA sequences for 135 species from 119 genera (5342 nucleotides). The ancestral pollination interaction in Araceae was recon- structed as probably rewarding albeit with low confidence because information is available for only 56 of the 120–130 genera. Bayesian stochastic trait mapping showed that spadix zonation, presence of an appendix, and flower sexuality were correlated with pollination interaction type. In the Araceae, having unisexual flowers appears to have provided the morphological precon- dition for the evolution of traps. Compared with the frequency of shifts between deceptive and rewarding pollination systems in orchids, our results indicate less lability in the Araceae, probably because of morphologically and sexually more specialized inflorescences. -

Caladium Genetics and Breeding: Recent Advances
® Floriculture and Ornamental Biotechnology ©2012 Global Science Books Caladium Genetics and Breeding: Recent Advances Zhanao Deng University of Florida/IFAS, Environmental Horticulture Department, Gulf Coast Research and Education Center, 14625 County Road 672, Wimauma, FL 33598, USA Corresponding author : [email protected] ABSTRACT Caladiums are important ornamental aroids; they are valued for their colourful and variably-shaped leaves. Numerous advances have been made in recent decades in caladium breeding and genetic studies. Techniques have been developed to increase flower production, store pollen, and maintain seed viability. Sources of genetic resistance have been identified for important diseases and pests (such as Fusarium tuber rot, Pythium root rot, bacterial blight, and root-knot nematodes) and abiotic stress factors including chilling injury. Mode of inheritance for important foliar traits has been elucidated through analysis of trait segregation in progeny populations. Caladiums have evolved three alleles at one locus that control colour of leaf main veins (red, white or green) and two co-dominant alleles at an independent locus that determine leaf shapes (fancy, lance, or strap). Gene loci for leaf spotting and blotching are both simply inherited but tightly linked to green veins. In vitro culture and plant regeneration were successful with several types of tissues/organs through somatic embryogenesis and/or organogenesis. Shoot-tip culture has been used to eliminate viral and fungal pathogens and invigorate planting stock; protoplasts isolated from leaf callus regenerated into whole plants; foreign genes from maize or humans have been introduced into caladium through Agrobacterium co-cultivation. Molecular markers, including highly specific and informative SSRs, have been developed and applied to caladium to distinguish cultivars, assess genetic diversity, and analyze genetic relationships. -
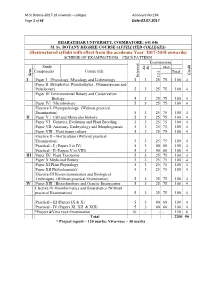
Restructured Syllabi with Effect from the Academic Year 2017-2018 Onwards
M.Sc Botany-2017-18 onwards – colleges Annexure No:19A Page 1 of 43 Date:03.07.2017 BHARATHIAR UNIVERSITY, COIMBATORE: 641 046 M. Sc. BOTANY DEGREE COURSE (AFFILIATED COLLEGES) (Restructured syllabi with effect from the academic Year 2017-2018 onwards) SCHEME OF EXAMINATIONS – CBCS PATTERN Examinations Study Mark Hrs. Dur. Components Course title Total Credit Sem. CIA Ins. hrs/week I Paper I Phycology, Mycology and Lichenology 5 3 25 75 100 4 Paper II Bryophytes, Pteridophytes , Gymnosperms and Paleobotany 5 3 25 75 100 4 Paper III Environmental Botany and Conservation Biology 5 3 25 75 100 4 Paper IV Microbiology 5 3 25 75 100 4 Elective I- Phytopathology (Without practical Examination) 5 3 25 75 100 4 II Paper V Cell and Molecular biology 5 3 25 75 100 4 Paper VI Genetics, Evolution and Plant Breeding 5 3 25 75 100 4 5 Paper VII Anatomy, Embryology and Morphogenesis 5 3 25 75 100 4 Paper VIII Plant tissue culture 5 3 25 75 100 4 Elective II – Horticulture (Without practical Examination) 5 3 25 75 100 4 Practical - I (Papers I to IV) 5 3 40 60 100 4 Practical - II (Papers V to VIII) 5 3 40 60 100 4 III Paper IX Plant Taxonomy 5 3 25 75 100 4 Paper X Medicinal Botany 5 3 25 75 100 4 Paper XI Plant Physiology 5 3 25 75 100 4 Paper XII Phytochemistry 5 3 25 75 100 4 Elective-III Bioinstrumentation and Biological Techniques (Without practical Examination) 5 3 25 75 100 4 IV Paper XIII Biotechnology and Genetic Engineering 5 3 25 75 100 4 Elective IV-Bioinformatics and Biostatistics (Without practical Examination) 5 3 25 75 100 4 Practical - III (Papers IX & X) 5 3 40 60 100 4 Practical - IV (Papers XI, XII & XIII) 5 3 40 60 100 4 *Project &Viva voce Examination 10 - - - 150 6 Total 2250 90 * Project report – 120 marks; Viva-voce – 30 marks M.Sc Botany-2017-18 onwards – colleges Annexure No:19A Page 2 of 43 Date:03.07.2017 Method of implementation and evaluation of Project Based on the strength, students will be allotted to staff members by lot in the first week after reopening the college. -

Natural Hybridization – Recombination – an Ever-Ongoing Process
THAI FOREST BULL., BOT. 47(1): 19–28. 2019. DOI https://doi.org/10.20531/tfb.2019.47.1.05 Natural hybridization – recombination – an ever-ongoing process NIELS JACOBSEN1,* & MARIAN ØRGAARD1 ABSTRACT Exemplified by studies of the SE Asian genusCryptocoryne (Araceae) we provide evidence that: 1) interspecific hybridization is an ever- ongoing process, and introgression and gene exchange takes place whenever physically possible throughout the region; 2) artificial hybridization experiments confirm that wide crosses are possible in a large number of cases; 3) rivers and streams provide numerous, diverse habitats for Cryptocoryne diaspores to settle in; 4) the changes in habitats caused by recurrent glaciations resulting in numerous splitting and merging of populations facilitates hybridization and segregation of subsequent generations; 5) hybridization is a major driving element in speciation; 6) populations are the units and stepping stones in evolution – not the species. KEYWORDS: Araceae, Chromosome numbers, Cryptocoryne, hybridization, evolution. Accepted for publication: 11 January 2019. Published online: 14 February 2019 INTRODUCTION Chiang Khan, Loei Province, hosts a number of morphologically distinct but genetically closely The completion of Cryptocoryne Fisch. ex related hybrids (Idei et al., 2017). Wydler for the Araceae volume for Flora of Thailand stands as a milestone after several years of research in this biological unique and complex genus (Boyce THE GENUS CRYPTOCORYNE et al., 2012). Numerous cultivation experiments, The genus -

History and Current Status of Systematic Research with Araceae
HISTORY AND CURRENT STATUS OF SYSTEMATIC RESEARCH WITH ARACEAE Thomas B. Croat Missouri Botanical Garden P. O. Box 299 St. Louis, MO 63166 U.S.A. Note: This paper, originally published in Aroideana Vol. 21, pp. 26–145 in 1998, is periodically updated onto the IAS web page with current additions. Any mistakes, proposed changes, or new publications that deal with the systematics of Araceae should be brought to my attention. Mail to me at the address listed above, or e-mail me at [email protected]. Last revised November 2004 INTRODUCTION The history of systematic work with Araceae has been previously covered by Nicolson (1987b), and was the subject of a chapter in the Genera of Araceae by Mayo, Bogner & Boyce (1997) and in Curtis's Botanical Magazine new series (Mayo et al., 1995). In addition to covering many of the principal players in the field of aroid research, Nicolson's paper dealt with the evolution of family concepts and gave a comparison of the then current modern systems of classification. The papers by Mayo, Bogner and Boyce were more comprehensive in scope than that of Nicolson, but still did not cover in great detail many of the participants in Araceae research. In contrast, this paper will cover all systematic and floristic work that deals with Araceae, which is known to me. It will not, in general, deal with agronomic papers on Araceae such as the rich literature on taro and its cultivation, nor will it deal with smaller papers of a technical nature or those dealing with pollination biology. -

A Review of the Aroid Tribe Caladieae with the Description of Three New Species of Caladium and Seven New Species of Syngonium (Araceae)
A Review of the Aroid Tribe Caladieae with the Description of Three New Species of Caladium and Seven New Species of Syngonium (Araceae) Thomas B. Croat,1 Xavier Delannay,2 Orlando O. Ortiz,3 and Pedro Diaz Jim´enez4 1 P. A. Schulze Curator, Missouri Botanical Garden, 4344 Shaw Blvd., St. Louis, Missouri 63110, U.S.A. [email protected] 2 Volunteer Research Associate, Missouri Botanical Garden, 4344 Shaw Blvd., St. Louis, Missouri 63110, U.S.A. 3 Herbario PMA & Programa de Maestr´ıa en Ciencias Biol´ogicas, Universidad de Panam´a, Estafeta Universitaria, Panama City, Panama. 4 Centro de Investigaciones Tropicales, Universidad Veracruzana, Xalapa, Veracruz, Mexico. ABSTRACT. A review of the aroid tribe Caladieae is that summarized our knowledge of the group, described presented, and three new species of Caladium Vent. new species, and discovered important distinctions and seven new species of Syngonium Schott are de- between Caladium and Xanthosoma, markedly that scribed and illustrated. Two species, C. picturatum K. differences occur in the display of their pollen. Pollen Koch & C. D. Bouch´e and C. steudnerifolium Engl., grains are borne in tetrads in Xanthosoma and solitarily previously considered to be synonymous with C. bicolor in Caladium. Madison (1981) provided a key to the six (Aiton) Vent., are fully redescribed. New species of genera considered to be Caladieae at that time, namely Caladium are C. cortesae Croat & E. G. Gonç., C. Aphyllarum S. Moore, Caladium, Chlorospatha, Jasa- palaciosii Croat & L. P. Hannon, and C. stevensonii rum, Scaphispatha, and Xanthosoma. Aphyllarum has Croat & Delannay. New species of Syngonium are S. -

Total Phenolics, Antioxidant, Antibacterial and Cytotoxic Activity Studies of Ethanolic Extracts Arisarum Vulgare O.Targ.Tozz
Int. J. Sec. Metabolite, Vol. 4, Issue 2 (2017) pp. 114-122 Research Article ISSN: 2148-6905 online Journal homepage: http://www.ijate.net/index.php/ijsm Total Phenolics, Antioxidant, Antibacterial and Cytotoxic Activity Studies of Ethanolic Extracts Arisarum vulgare O.Targ.Tozz. and Dracunculus vulgaris Schott. Çiğdem AYDIN1,*, Cennet ÖZAY1, Olcay DÜŞEN1, Ramazan MAMMADOV1, Figen ORHAN1 1Pamukkale University, Science & Art Faculty, Department of Biology, 20070, Denizli, Turkey Received: 08 February 2017 - Revised: 20 March 2017 - Accepted: 18 April 2017 Abstract: The present study reports the antioxidant, total phenolics, antibacterial and cytotoxic activities of Arisarum vulgare and Dracunculus vulgaris of aerial and under ground ethanol extracts. The antioxidant activity was determined by DPPH radical scavenging activity and β-carotene linoleic acid assays. Folin-Ciocalteu assay was used to detect total phenolic contents in the extract. In vitro cytotoxic activity was determined by the Brine shrimp lethality test. Antibacterial activity was investigated with the microdilution method. As a result, under ground parts of the extract showed higher antioxidant activity than aerial parts of the extract. Total phenolics ranged from 14.5±2.02 to 53.4±2.01 mg GAE/g, and the antioxidant activity according to the β-carotene/linoleic acid assay ranged from 70.76±1.08 % to 85.43±1.05 and according to the DPPH assay IC50 values ranged from 0.089±1.02 to 1.095±1.07mg/ml. The A. vulgare under ground extract was tested against 2 and showed a good antibacterial activity at a concentration of <50 g/mL Minimum inhibitory concentration (MIC) values for the bacteria S.aureus. -

Hardy Aroids in the Garden Judy Glattstein
Hardy Aroids in the Garden Judy Glattstein Though not showy plants and with only a modest following among plant lovers, the hardy aroids are interesting, display many virtues in cultivation, and attract "a different class of gardeners" The Arum Family, or Araceae, consists of tion, especially in the United States. They about fifteen genera, most of them tropi- are, therefore, unusual and have the cal but of wide distribution. Some of the appeal of novelty. tropical members of the family have long Aroids contain a bitter substance, calci- been under cultivation, especially in east- um oxalate, and are little bothered by ern Asia and the Pacific Islands. Taro pests. Slugs, mice, rabbits, and deer find (Colocasia esculenta) and several species them decidedly unpalatable. When aroids of Xanthosoma (yautia), for example, are are used for food, the calcium oxalate grown for their edible tubers as staple first must be destroyed by heat. Garden- sources of starch. Other tropical species ers should be careful to wash their hands are handsome foliage plants used in the after handling berries or a bruised tuber. temperate zones for summer bedding (Ca- Once, after cleaning Aris~ma seeds, I ladium) or as houseplants (Aglaonema, inadvertently touched my mouth. The Dieffenbachia, Monstera, Philodendron). resulting unpleasant tingling and numb- Others are used by florists as cut flowers ness took several hours to wear off. (Anthurium, Calla~.I. My garden in Wilton, Connecticut, is Some members of the family are hardy, shaded by mature white oaks (Quercus notably Aris~ma, Arisarum, Arum, Lysi- alba). Understory trees are dogwood chiton, and Symplocarpus.