HABITAT USE and NESTING ECOLOGY of RING-NECKED PHEASANT (Phasianus Colchicus)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Keeping Pheasants
KKKEEEEEEPPPIIINNNGGG PPPHHHEEEAAASSSAAANNNTTTSSS AAASSS AAA HHHOOOBBBBBBYYY Text and photos: Jan Willem Schrijvers Photo above: Siamese Fireback pheasant male(Lophura diardi). ORIGIN AND LIFESTYLE OF THE WILD PHEASANT Pheasants are wild gallinaceous birds all originating in Asia. One exception is the Congo peacock from Africa. (The pheasants also include game fowl and peacocks.) Each species has its own characteristics and life habits. There are species that live in tropical rain forests, but there are species that live in the mountains, on cold plains. This is something to take in account when housing our pheasants (with or without a night coop, with or without heating). Most species live in and around the mountains with a woodland vegetation, where they can find lots of berries, greens and seeds. It is of utmost importance to first consider the habits and living conditions of the species you want to keep. Only with proper housing will these beautiful birds show to their fullest, and reproduce. PHEASANTS IN AVIARIES In the past there were some "pheasant farms" which mainly bred the common pheasant (also known as ring-neck pheasant). The birds on these farms were bred for sport hunting. They were released en masse in the autumn in the fields and other areas to be shot for the sport. Some escaped from the hunters and that is why today we can see wild pheasants roam our fields and woods. These pheasants have also succeeded in reproducing themselves, even though they are not native birds. The birds are also kept for their colourful feathers. Each year I see Prince Carnival walk again with the beautiful feathers of the Reeves's Pheasant at his cocked hat. -

Sehr Geehrte Damen Und Herren! Werte Mitbürgerinnen Und Mitbürger! Bei Der Gemeinderatssitzung Am 31
Sehr geehrte Damen und Herren! Werte Mitbürgerinnen und Mitbürger! Bei der Gemeinderatssitzung am 31. Okto in einem eigenen Termin am Freitag, den 14. ber 2013 wurden entscheidende Weichen für Februar 2014, um 19 Uhr im GH Holzer in die Zukunft unserer Gemeinde gestellt. formieren. Dabei wird auch ausgelotet, inwie Nach längeren Verhandlungen konnte der weit tatsächlich Interesse von Jugendlichen Vertrag mit dem Wohnbauträger „Austria“ unserer Gemeinde besteht – dies ist dann für abgeschlossen werden, und somit wird das den Wohnbauträger die Voraussetzung, ob „Betreute Wohnen“ bald Realität in unserer das Vorhaben realisiert wird. Gemeinde. Über die Wintermonate sollen Ich kann nur alle Jugendlichen, die gerne dann noch alle baurechtlichen Fakten abge ihren Start ins Leben in unserer Marktge schlossen werden – im Frühjahr wird gebaut. meinde Wullersdorf beginnen wollen, herzlich Ebenso kauften wir bei dieser Sitzung das einladen, von diesem Angebot Gebrauch zu restliche Areal von Herrn DI Daniel Brabenetz, machen! und beschlossen gleichzeitig auch einen Ver In diesem Sinne danke ich allen aktiven trag mit dem Wohnbauträger „Waldviertel“, Kräften in unserer Gemeinde für die stets welcher uns auch ein so genanntes „Junges gute Zusammenarbeit und wünsche Ihnen Wohnen“ ermöglichen soll. Diese Wohnan zum bevorstehenden Weihnachtsfest und lage würde dann unter anderem auf diesem dem Jahreswechsel alles erdenklich Gute, Areal errichtet und soll als „Startwohnungen“ vor allem Gesundheit. dienen. Ihr Richard HOGL e.h. Über das genaue Prozedere, die Voraus Bürgermeister setzungen und Bestimmungen werden wir (0676) 401 42 67 Der Dorftrommler Dezember 2013 Finanzielle Unterstützungen Insgesamt erhielt unsere Gemeinde hier mit im letzten Quartal 2013 Förderungen in durch das Land Niederösterreich der Höhe von R 48.338,65, plus einer Zusa R Die NÖ Landesregierung hat in Ihrer Sit ge über 35.000,–, wofür ich als Bürgermei zung vom 1. -

A Study of Food and Feeding Habits of Blue Peafowl, Pavo Cristatus Linnaeus, 1758 in District Kurukshetra, Haryana (India)
International Journal of Research Studies in Biosciences (IJRSB) Volume 2, Issue 6, July 2014, PP 11-16 ISSN 2349-0357 (Print) & ISSN 2349-0365 (Online) www.arcjournals.org A Study of Food and Feeding Habits of Blue Peafowl, Pavo Cristatus Linnaeus, 1758 in District Kurukshetra, Haryana (India) Girish Chopra, Tarsem Kumar Department of Zoology, Kurukshetra University, Kurukshetra-136119 (INDIA) [email protected] Summary: Present study was conducted to determine the food and feeding habits of blue peafowl in three study sites, namely, Saraswati plantation wildlife sanctuary (SPWS), Bir Sonti Reserve Forest (BSRF), and Jhrouli Kalan village (JKAL). Point count method (Blondel et al., 1981) was followed during periodic fortnightly visits to all the three selected study sites. The peafowls were observed to feed on flowers, fruits, leaves of 11, 8 and 8 plant species respectively. These were sighted to feed on Brassica compestris (flowers, leaves), Trifolium alexandarium (flowers, leaves), Triticum aestivum (flowers, leaves, fruits), Oryza sativa (flowers, leaves, fruits), Chenopodium album (flowers, leaves, fruits), Parthenium histerophoresus (flowers, leaves), Pisum sativum (flowers, leaves, fruits), Cicer arientum (flowers, leaves, fruits), Pyrus pyrifolia (flowers, fruits), Ficus benghalensis (flowers, fruits), Ficus rumphii (flowers, fruits). They were also observed feeding on insects in all three study sites and on remains of the snake bodies at the BSRF and JKAL study site. The findings revealed that the Indian peafowl, on one hand, functions as a predator of agricultural pests but, on the other hand, is itself a pest on agricultural crops. Keywords: Blue peafowl, Food, Feeding Habits, Herbs, Shrubs, Trees. 1. INTRODUCTION Birds are warm-blooded, bipedal, oviparous vertebrates characterized by bony beak, pneumatic bones, feathers and wings. -

Mittelalter.Pdf
Herrschaftsbildung und wirtschaftlicher Mißerfolg – Guntersdorf im späten Mittelalter (1258‐1476) Von Markus Jeitler Einleitung Die Geschichte unseres Ortes läßt sich in den Jahrzehnten nach seiner ersten 1 schriftlichen Nennung (1108 bzw. 1141)TPD DPT mangels erhaltener Quellen so gut wie nicht erfassen. Dies liegt vermutlich daran, daß ein großer Teil des Melker Stiftarchivs im Jahre 1297 einer Feuersbrunst zum Opfer fiel und somit wohl auch für unsere Fragestellungen relevante Stücke verloren gingen. Aus späteren Hinweisen und Überlieferungen kann man aber feststellen, daß das Stift Melk bis 1538 weiterhin die Lehenshoheit in Guntersdorf innehatte und seine Güter jeweils an Lehensträger ausgab, was noch auszuführen sein wird. An bisherigen Zusammenfassungen zur mittelalterlichen und neuzeitlichen Geschichte Guntersdorfs seien ein Beitrag von 2 Joseph Feil,TPD DPT dem der entsprechende Artikel in der „Topographie von 3 Niederösterreich“TPD DPT im wesentlichen folgt, und eine Zusammenstellung von Gertrude 4 Langer‐Ostrawsky im Heimatbuch des Bezirkes Hollabrunn genannt.TPD DPT Die Zeit der Übernahme der Herrschaft durch die Familie der Wallseer wird schließlich durch 5 Karel Hruza exemplarisch behandelt;TPD DPT Ignaz Franz Keiblinger geht in seinem umfangreichen Werk über die Geschichte des Stiftes Melk im Rahmen der 6 Darstellung der Pfarre Wullersdorf ebenfalls auf Guntersdorf ein.TPD DPT Darüber hinaus konnten weitere Hinweise gefunden werden, die das Gesamtbild abrunden bzw. für 7 Großnondorf neue Erkenntnisse bringen werden.TPD DPT Besitzungen zu Guntersdorf. Die ersten Nachrichten, die die Geschichte unseres Ortes im 13. Jahrhundert beleuchten, betreffen das Stift Heiligenkreuz. König Ottokar II. Přemysl bestätigt im Jahre 1258 die Schenkung des Albero von Leis über vier Mansen, aus seinem 8 Eigengut, die er nun zu seinem Seelenheil dem Stift übergibt.TPD DPT Diese Schenkung wird am 24. -

Klima- Und Energie-Modellregionen
Klima- und Energie-Modellregion : KLIMA- UND ENERGIEMODELLREGION PULKAUTAL Bericht der Umsetzungsphase Weiterführungsphase I Weiterführungsphase II Weiterführungsphase III Zwischenbericht Endbericht Inhaltsverzeichnis: 1. Fact-Sheet zur Klima- und Energie-Modellregion 2. Zielsetzung 3. Eingebundene Akteursgruppen 4. Aktivitätenbericht 5. Best Practice Beispiel der Umsetzung 1. Fact-Sheet zur Klima- und Energie-Modellregion Facts zur Klima- und Energie-Modellregion Name der Klima- und Energiemodellregion (KEM): Klima- und Energiemodellregion Pulkautal (Offizielle Regionsbezeichnung) Geschäftszahl der KEM B287567 Trägerorganisation, Rechtsform Initiative Pulkautal – Verein zur Entwicklungsförde- rung der Gemeinden des Gerichtbezirkes Haugsdorf Deckt sich die Abgrenzung und Bezeichnung der X Ja KEM mit einem bereits etablierten Regionsbegriff (j/n)? Initiative Pulkautal Falls ja, bitte Regionsbezeichnung anführen: Facts zur Klima- und Energiemodellregion: - Anzahl der Gemeinden: Die KEM umfasst insgesamt 6 Gemeinden - Anzahl der Einwohner/innen: 6.543 Einwohnern in der KEM - geografische Beschreibung (max. 400 Zeichen) Die Region Pulkautal liegt im nördlichen Weinviertel in Niederösterreich im politischen Bezirk Hol- labrunn. Das Pulkautal grenzt direkt an die Tsche- chische Republik. Namensgeber des Tales ist der Fluss Pulkau. Die Region liegt zwischen zwei Bal-lungszentren, ca. 80 Kilometer nord-westlich von Wien an der Grenze zu Tschechien, nur 15 Kilometer von Znaim und ca. 80 Kilometer von Brünn entfernt. Modellregions-Manager/in (MRM) -

Hybridization & Zoogeographic Patterns in Pheasants
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Paul Johnsgard Collection Papers in the Biological Sciences 1983 Hybridization & Zoogeographic Patterns in Pheasants Paul A. Johnsgard University of Nebraska-Lincoln, [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/johnsgard Part of the Ornithology Commons Johnsgard, Paul A., "Hybridization & Zoogeographic Patterns in Pheasants" (1983). Paul Johnsgard Collection. 17. https://digitalcommons.unl.edu/johnsgard/17 This Article is brought to you for free and open access by the Papers in the Biological Sciences at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Paul Johnsgard Collection by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. HYBRIDIZATION & ZOOGEOGRAPHIC PATTERNS IN PHEASANTS PAUL A. JOHNSGARD The purpose of this paper is to infonn members of the W.P.A. of an unusual scientific use of the extent and significance of hybridization among pheasants (tribe Phasianini in the proposed classification of Johnsgard~ 1973). This has occasionally occurred naturally, as for example between such locally sympatric species pairs as the kalij (Lophura leucol11elana) and the silver pheasant (L. nycthelnera), but usually occurs "'accidentally" in captive birds, especially in the absence of conspecific mates. Rarely has it been specifically planned for scientific purposes, such as for obtaining genetic, morphological, or biochemical information on hybrid haemoglobins (Brush. 1967), trans ferins (Crozier, 1967), or immunoelectrophoretic comparisons of blood sera (Sato, Ishi and HiraI, 1967). The literature has been summarized by Gray (1958), Delacour (1977), and Rutgers and Norris (1970). Some of these alleged hybrids, especially those not involving other Galliformes, were inadequately doculnented, and in a few cases such as a supposed hybrid between domestic fowl (Gallus gal/us) and the lyrebird (Menura novaehollandiae) can be discounted. -

Europe's Huntable Birds a Review of Status and Conservation Priorities
FACE - EUROPEAN FEDERATIONEurope’s FOR Huntable HUNTING Birds A Review AND CONSERVATIONof Status and Conservation Priorities Europe’s Huntable Birds A Review of Status and Conservation Priorities December 2020 1 European Federation for Hunting and Conservation (FACE) Established in 1977, FACE represents the interests of Europe’s 7 million hunters, as an international non-profit-making non-governmental organisation. Its members are comprised of the national hunters’ associations from 37 European countries including the EU-27. FACE upholds the principle of sustainable use and in this regard its members have a deep interest in the conservation and improvement of the quality of the European environment. See: www.face.eu Reference Sibille S., Griffin, C. and Scallan, D. (2020) Europe’s Huntable Birds: A Review of Status and Conservation Priorities. European Federation for Hunting and Conservation (FACE). https://www.face.eu/ 2 Europe’s Huntable Birds A Review of Status and Conservation Priorities Executive summary Context Non-Annex species show the highest proportion of ‘secure’ status and the lowest of ‘threatened’ status. Taking all wild birds into account, The EU State of Nature report (2020) provides results of the national the situation has deteriorated from the 2008-2012 to the 2013-2018 reporting under the Birds and Habitats directives (2013 to 2018), and a assessments. wider assessment of Europe’s biodiversity. For FACE, the findings are of key importance as they provide a timely health check on the status of In the State of Nature report (2020), ‘agriculture’ is the most frequently huntable birds listed in Annex II of the Birds Directive. -
Phylogeography of the Common Pheasant Phasianus Colchinus
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/311941340 Phylogeography of the Common Pheasant Phasianus colchinus Article in Ibis · February 2017 DOI: 10.1111/ibi.12455 CITATION READS 1 156 5 authors, including: Nasrin Kayvanfar Mansour Aliabadian Ferdowsi University Of Mashhad Ferdowsi University Of Mashhad 7 PUBLICATIONS 39 CITATIONS 178 PUBLICATIONS 529 CITATIONS SEE PROFILE SEE PROFILE Zheng-Wang Zhang Yang Liu Beijing Normal University Sun Yat-Sen University 143 PUBLICATIONS 827 CITATIONS 53 PUBLICATIONS 286 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Using of microvertebrate remains in reconstruction of late quaternary (Holocene) paleoclimate, Eastern Iran View project Spatial distribution and composition of aliphatic hydrocarbons, polycyclic aromatic hydrocarbons and hopanes in superficial sediments of the coral reefs of the Persian Gulf, Iran View project All content following this page was uploaded by Yang Liu on 05 March 2017. The user has requested enhancement of the downloaded file. All in-text references underlined in blue are added to the original document and are linked to publications on ResearchGate, letting you access and read them immediately. Ibis (2016), doi: 10.1111/ibi.12455 Phylogeography of the Common Pheasant Phasianus colchicus NASRIN KAYVANFAR,1 MANSOUR ALIABADIAN,1,2* XIAOJU NIU,3 ZHENGWANG ZHANG3 & YANG LIU4* 1Department of Biology, Faculty of Science, Ferdowsi University -

Phylogenetic Relationships of the Phasianidae Reveals Possible Non-Pheasant Taxa
Journal of Heredity 2003:94(6):472–489 Ó 2003 The American Genetic Association DOI: 10.1093/jhered/esg092 Phylogenetic Relationships of the Phasianidae Reveals Possible Non-Pheasant Taxa K. L. BUSH AND C. STROBECK From the Department of Biological Sciences, University of Alberta–Edmonton, Edmonton, Alberta T6G 2E9, Canada. Address correspondence to Krista Bush at the address above, or e-mail: [email protected]. Abstract The phylogenetic relationships of 21 pheasant and 6 non-pheasant species were determined using nucleotide sequences from the mitochondrial cytochrome b gene. Maximum parsimony and maximum likelihood analysis were used to try to resolve the phylogenetic relationships within Phasianidae. Both the degree of resolution and strength of support are improved over previous studies due to the testing of a number of species from multiple pheasant genera, but several major ambiguities persist. Polyplectron bicalcaratum (grey peacock pheasant) is shown not to be a pheasant. Alternatively, it appears ancestral to either the partridges or peafowl. Pucrasia macrolopha macrolopha (koklass) and Gallus gallus (red jungle fowl) both emerge as non-pheasant genera. Monophyly of the pheasant group is challenged if Pucrasia macrolopha macrolopha and Gallus gallus are considered to be pheasants. The placement of Catreus wallichii (cheer) within the pheasants also remains undetermined, as does the cause for the great sequence divergence in Chrysolophus pictus obscurus (black-throated golden). These results suggest that alterations in taxonomic -
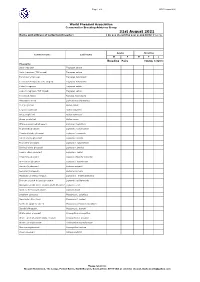
31St August 2021 Name and Address of Collection/Breeder: Do You Closed Ring Your Young Birds? Yes / No
Page 1 of 3 WPA Census 2021 World Pheasant Association Conservation Breeding Advisory Group 31st August 2021 Name and address of collection/breeder: Do you closed ring your young birds? Yes / No Adults Juveniles Common name Latin name M F M F ? Breeding Pairs YOUNG 12 MTH+ Pheasants Satyr tragopan Tragopan satyra Satyr tragopan (TRS ringed) Tragopan satyra Temminck's tragopan Tragopan temminckii Temminck's tragopan (TRT ringed) Tragopan temminckii Cabot's tragopan Tragopan caboti Cabot's tragopan (TRT ringed) Tragopan caboti Koklass pheasant Pucrasia macrolopha Himalayan monal Lophophorus impeyanus Red junglefowl Gallus gallus Ceylon junglefowl Gallus lafayettei Grey junglefowl Gallus sonneratii Green junglefowl Gallus varius White-crested kalij pheasant Lophura l. hamiltoni Nepal Kalij pheasant Lophura l. leucomelana Crawfurd's kalij pheasant Lophura l. crawfurdi Lineated kalij pheasant Lophura l. lineata True silver pheasant Lophura n. nycthemera Berlioz’s silver pheasant Lophura n. berliozi Lewis’s silver pheasant Lophura n. lewisi Edwards's pheasant Lophura edwardsi edwardsi Vietnamese pheasant Lophura e. hatinhensis Swinhoe's pheasant Lophura swinhoii Salvadori's pheasant Lophura inornata Malaysian crestless fireback Lophura e. erythrophthalma Bornean crested fireback pheasant Lophura i. ignita/nobilis Malaysian crestless fireback/Vieillot's Pheasant Lophura i. rufa Siamese fireback pheasant Lophura diardi Southern Cavcasus Phasianus C. colchicus Manchurian Ring Neck Phasianus C. pallasi Northern Japanese Green Phasianus versicolor -
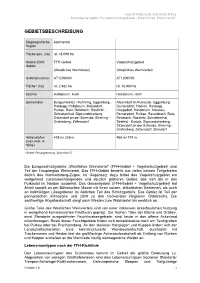
Gebietsbeschreibung
HAUPTREGION WEINVIERTEL Managementplan Europaschutzgebiete „Westliches Weinviertel“ GEBIETSBESCHREIBUNG Biogeografische kontinental Region Fläche ges. (ha) rd. 18.099 ha Natura 2000- FFH-Gebiet Vogelschutzgebiet Gebiet (Westliches Weinviertel) (Westliches Weinviertel) Gebietsnummer AT1209A00 AT1209V00 Fläche* (ha) rd. 2.982 ha rd. 16.904 ha Bezirke Hollabrunn, Horn Hollabrunn, Horn Gemeinden Burgschleinitz - Kühnring, Eggenburg, Alberndorf im Pulkautal, Eggenburg, Hardegg, Hollabrunn, Meiseldorf, Guntersdorf, Hadres, Hardegg, Pulkau, Retz, Retzbach, Röschitz, Haugsdorf, Hollabrunn, Maissau, Schrattenthal, Sigmundsherberg, Pernersdorf, Pulkau, Ravelsbach, Retz, Sitzendorf an der Schmida, Straning - Retzbach, Röschitz, Schrattenthal, Grafenberg, Zellerndorf Seefeld - Kadolz, Sigmundsherberg, Sitzendorf an der Schmida, Straning - Grafenberg, Zellerndorf, Ziersdorf Höhenstufen 438 m/ 226 m 468 m/ 191 m (max./min. m Höhe) * Quelle: Feinabgrenzung, Stand Mai 07 Die Europaschutzgebiete „Westliches Weinviertel” (FFH-Gebiet + Vogelschutzgebiet) sind Teil der Hauptregion Weinviertel. Das FFH-Gebiet besteht aus vielen kleinen Teilgebieten östlich des Manhartsberg-Zuges. Im Gegensatz dazu bildet das Vogelschutzgebiet ein weitgehend zusammenhängendes und deutlich größeres Gebiet, das sich bis in das Pulkautal im Norden ausdehnt. Das Gesamtgebiet (FFH-Gebiet + Vogelschutzgebiet) hat Anteil sowohl an der Böhmischen Masse mit ihren sauren, silikatischen Gesteinen, als auch an kalkhältigen Lössgebieten im östlichen Teil des Schutzgebiets. Das Gebiet ist -

Patterns of Success in Game Bird Introductions in the United States
February 18, 2016 1 Patterns of Success in Game Bird Introductions in the United States 2 3 4 by 5 6 7 8 1Michael P. Moulton, 2Wendell P. Cropper, Jr., 1Andrew J. Broz, 2Salvador A. 9 Gezan 10 11 1Department of Wildlife Ecology and Conservation; University of Florida; PO 12 Box 110430; Gainesville, FL 32611-0430; <[email protected]> 13 2School of Forest Resources and Conservation; University of Florida; PO Box 14 110410; Gainesville, FL 32611-0410; <[email protected]> 15 16 "There is no safety in numbers, or in anything else." James Thurber, The Fairly 17 Intelligent Fly, The New Yorker, February 4, 1939. 18 19 20 1 PeerJ PrePrints | https://doi.org/10.7287/peerj.preprints.1765v1 | CC-BY 4.0 Open Access | rec: 20 Feb 2016, publ: 20 Feb 2016 February 18, 2016 21 Abstract 22 Better predictions of the success of species’ introductions require careful 23 evaluation of the relative importance of at least three kinds of factors: species 24 characteristics, characteristics of the site of introduction, and event-level factors 25 such as propagule pressure. Historical records of bird introductions provide a 26 unique method for addressing the relative importance of these factors. We compiled 27 a list of introductions of 17 Phasianid species released in the USA during the Foreign 28 Game Investigation Program (FGIP). These records indicate that releases for some 29 Phasianid species in the USA continued long after establishment. For many of the 13 30 species that always failed, even numerous releases and large numbers of individuals 31 per release were not enough for successful establishment, yet several of these 32 species were successfully introduced elsewhere.