Wetland Birds in the Recent Fossil Record of Britain and Northwest Europe John R
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ardea Cinerea (Grey Heron) Family: Ardeidae (Herons and Egrets) Order: Ciconiiformes (Storks, Herons and Ibises) Class: Aves (Birds)
UWU The Online Guide to the Animals of Trinidad and Tobago Behaviour Ardea cinerea (Grey Heron) Family: Ardeidae (Herons and Egrets) Order: Ciconiiformes (Storks, Herons and Ibises) Class: Aves (Birds) Fig. 1. Grey heron, Ardea cinerea. [http://www.google.tt/imgres?imgurl=http://www.bbc.co.uk/lancashire/content/images/2006/06/15/grey_heron, downloaded 14 November 2012] TRAITS. Grey herons are large birds that can be 90-100cm tall and an adult could weigh in at approximately 1.5 kg. They are identified by their long necks and very powerful dagger like bills (Briffett 1992). They have grey plumage with long black head plumes and their neck is white with black stripes on the front. In adults the forehead sides of the head and the centre of the crown are white. In flight the neck is folded back with the wings bowed and the flight feathers are black. Each gender looks alike except for the fact that females have shorter heads (Seng and Gardner 1997). The juvenile is greyer without black markings on the head and breast. They usually live long with a life span of 15-24 years. ECOLOGY. The grey heron is found in Europe, Asia and Africa, and has been recorded as an accidental visitor in Trinidad. Grey herons occur in many different habitat types including savannas, ponds, rivers, streams, lakes and temporary pools, coastal brackish water, wetlands, marsh and swamps. Their distribution may depend on the availability of shallow water (brackish, saline, fresh, flowing and standing) (Briffett 1992). They prefer areas with tall trees for nesting UWU The Online Guide to the Animals of Trinidad and Tobago Behaviour (arboreal rooster and nester) but if trees are unavailable, grey herons may roost in dense brush or undergrowth. -

The Grey Heron
Bird Life The Grey Heron t is quite likely that if someone points out a grey heron to you, I you will remember it the next time you see it. The grey heron is a tall bird, usually about 80cm to 1m in height and is common to inland waterways and coasts. Though the grey heron has a loud “fraank” call, it can most often be seen standing silently in shallow water with its long neck outstretched, watching the water for any sign of movement. The grey heron is usually found on its own, although some may feed close together. Their main food is fish, but they will take small mammals, insects, frogs and even young birds. Because of their habit of occasionally taking young birds, herons are not always popular and are often driven away from a feeding area by intensive mobbing. Mobbing is when smaller birds fly aggressively at their predator, in this case the heron, in order to defend their nests or their lives. Like all herons, grey herons breed in a colony called a heronry. They mostly nest in tall trees and bushes, but sometimes they nest on the ground or on ledge of rock by the sea. Nesting starts in February,when the birds perform elaborate displays and make noisy callings. They lay between 3-5 greenish-blue eggs, often stained white by the birds’ droppings. Once hatched, the young © Illustration: Audrey Murphy make continuous squawking noises as they wait to be fed by their parents. And though it doesn’t sound too pleasant, the parent Latin Name: Ardea cinerea swallows the food and brings it up again at the nest, where the Irish Name: Corr réisc young put their bills right inside their parents mouth in order to Colour: Grey back, white head and retrieve it! neck, with a black crest on head. -

Flamingo ABOUT the GROUP
Flamingo ABOUT THE GROUP Bulletin of the IUCN-SSC/Wetlands International The Flamingo Specialist Group (FSG) was established in 1978 at Tour du Valat in France, under the leadership of Dr. Alan Johnson, who coordinated the group until 2004 (see profile at www.wetlands.org/networks/Profiles/January.htm). Currently, the group is FLAMINGO SPECIALIST GROUP coordinated from the Wildfowl & Wetlands Trust at Slimbridge, UK, as part of the IUCN- SSC/Wetlands International Waterbird Network. The FSG is a global network of flamingo specialists (both scientists and non- scientists) concerned with the study, monitoring, management and conservation of the world’s six flamingo species populations. Its role is to actively promote flamingo research and conservation worldwide by encouraging information exchange and cooperation amongst these specialists, and with other relevant organisations, particularly IUCN - SSC, Ramsar, WWF International and BirdLife International. FSG members include experts in both in-situ (wild) and ex-situ (captive) flamingo conservation, as well as in fields ranging from field surveys to breeding biology, diseases, tracking movements and data management. There are currently 165 members around the world, from India to Chile, and from France to South Africa. Further information about the FSG, its membership, the membership list serve, or this bulletin can be obtained from Brooks Childress at the address below. Chair Assistant Chair Dr. Brooks Childress Mr. Nigel Jarrett Wildfowl & Wetlands Trust Wildfowl & Wetlands Trust Slimbridge Slimbridge Glos. GL2 7BT, UK Glos. GL2 7BT, UK Tel: +44 (0)1453 860437 Tel: +44 (0)1453 891177 Fax: +44 (0)1453 860437 Fax: +44 (0)1453 890827 [email protected] [email protected] Eastern Hemisphere Chair Western Hemisphere Chair Dr. -
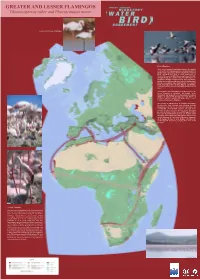
GREATER and LESSER FLAMINGOS Phoenicopterus Ruber and Phoeniconaias Minor
GREATER AND LESSER FLAMINGOS Phoenicopterus ruber and Phoeniconaias minor Greater and Lesser Flamingos © Cliff Buckton © P & H Harris Lesser Flamingo The Lesser Flamingo Phoeniconaias minor is the smallest of the world's five flamingo species. It occurs primarily in the Rift Valley lakes of East Africa with about 4 to 5 million birds estimated, but also in small populations in Namibia/Botswana (40,000), Mauritania/Senegal (15,400), Ethiopia (8,300). The alkaline lakes of the Rift Valley are the primary feeding areas for the East Africa population. During non-breeding periods these lakes often hold almost the entire population. Huge feeding flocks of 1-2 million birds frequently gather on lakes Bogoria and Nakuru, creating one of the most stunning wildlife spectacles in the world. Although it is still the most numerous of the five species, the Lesser Flamingo is classified as globally "near threatened" due primarily to its dependence on a limited number of unprotected breeding sites and threats of proposed soda-ash mining and hydro-electric power schemes on the main breeding lakes. The question of whether there is occasional interchange between the East African and southern African populations has yet to be resolved definitely, but considerable circumstantial evidence has now been assembled to show that East African Lesser Flamingos probably do fly to Botswana to breed during periods when the Lake Makgadikgadi Salt Pans are flooded. Their migration routes, flight range and stopover places (if any) are still unknown. It is now known that Lesser Flamingos do fly during the day, at great heights, well above the normal diurnal movement of eagles, their main aerial predator. -

Dalmatian Pelican Identification Manual Companion Document to the “Dalmatian Pelican Monitoring Manual”
Pelican Way of LIFE (LIFE18 NAT/NL/716) “Conservation of the Dalmatian Pelican along the Black Sea - Mediterranean Flyway” Dalmatian Pelican Identification Manual Companion document to the “Dalmatian Pelican Monitoring Manual” December 2020 Authored by: Commissioned by: Giorgos Catsadorakis and Olga Alexandrou, Society for the Protection of Prespa Rewilding Europe Dalmatian Pelican Identification Manual - Companion document to the “Dalmatian Pelican Monitoring Manual” December 2020 Authors: Giorgos Catsadorakis1 and Olga Alexandrou2 1,2Society for the Protection of Prespa, Agios Germanos, GR-53150, Prespa, Greece, [email protected] , [email protected] © Rewilding Europe Executed under the framework of “Conservation of the Dalmatian Pelican along the Black Sea - Mediterranean Flyway” (Pelican Way of LIFE; LIFE18 NAT/NL/716; https://life-pelicans.com/) project, financed by the LIFE programme of the European Union and Arcadia – a charitable fund of Lisbet Rausing and Peter Baldwin. Suggested citation: Society for the Protection of Prespa, 2020. Dalmatian pelican identification manual - Companion document to the “Dalmatian pelican monitoring manual”. Rewilding Europe. Produced within the framework of Pelican Way of LIFE project (LIFE18 NAT/NL/716). Keywords: Dalmatian pelican, identification, ageing, plumage, moulting patterns, sexing. Photo credits: Society for the Protection of Prespa, unless stated otherwise. Introduction Visual identification of the Dalmatian pelican seems like a straightforward task, yet in sites used by both species (the Dalmatian pelican and the great white pelican) identification can be tricky, especially when the observer is located some distance away from the birds. Hardly any useful material has been published on the sequence of moults and plumages of the Dalmatian pelican, and this gap often creates confusion and uncertainty about the accuracy of data in regards to ageing of Dalmatian pelicans, as well as frequently undermining confidence in census numbers. -

Flamingo Newsletter 17, 2009
ABOUT THE GROUP The Flamingo Specialist Group (FSG) is a global network of flamingo specialists (both scientists and non-scientists) concerned with the study, monitoring, management and conservation of the world’s six flamingo species populations. Its role is to actively promote flamingo research, conservation and education worldwide by encouraging information exchange and cooperation among these specialists, and with other relevant organisations, particularly the IUCN Species Survival Commission (SSC), the Ramsar Convention on Wetlands, the Convention on Conservation of Migratory Species (CMS), the African-Eurasian Migratory Waterbird Agreement (AEWA), and BirdLife International. The group is coordinated from the Wildfowl & Wetlands Trust, Slimbridge, UK, as part of the IUCN-SSC/Wetlands International Waterbird Network. FSG members include experts in both in-situ (wild) and ex-situ (captive) flamingo conservation, as well as in fields ranging from research surveys to breeding biology, infectious diseases, toxicology, movement tracking and data management. There are currently 286 members representing 206 organisations around the world, from India to Chile, and from France to South Africa. Further information about the FSG, its membership, the membership list serve, or this bulletin can be obtained from Brooks Childress at the address below. Chair Dr. Brooks Childress Wildfowl & Wetlands Trust Slimbridge Glos. GL2 7BT, UK Tel: +44 (0)1453 860437 Fax: +44 (0)1453 860437 [email protected] Eastern Hemisphere Chair Western Hemisphere Chair Dr. Arnaud Béchet Dr. Felicity Arengo Station biologique, Tour du Valat American Museum of Natural History Le Sambuc Central Park West at 79th Street 13200 Arles, France New York, NY 10024 USA Tel : +33 (0) 4 90 97 20 13 Tel: +1 212 313-7076 Fax : +33 (0) 4 90 97 20 19 Fax: +1 212 769-5292 [email protected] [email protected] Citation: Childress, B., Arengo, F. -

Comments on the Population Status of Chilean Flamingos at Lagoa Do Peixe National Park, Southern Brazil
Delfino and Aldana-Ardila. Flamingo 2020, pages: 21-26. Comments on the population status of Chilean flamingos at Lagoa do Peixe National Park, Southern Brazil Henrique Cardoso Delfino 1* & Oscar Maurício Aldana-Ardila 1 1 Universidade Federal do Rio Grande do Sul, Instituto de Biociências, Departamento de Zoologia, Programa de Pós-Graduação em Biologia Animal, Laboratório de Ecologia e Sistemática de Aves e Mamíferos Marinhos (LABSMAR). Av. Bento Gonçalves, 9500. CEP: 91509-900, Porto Alegre, RS, Brasil. *Corresponding author: [email protected] Abstract In South America, the Chilean flamingo (Phoenicopterus chilensis) is distributed from south of the Equator to southern Argentina, passing by the Brazilian coast. One of the locations where this species is present in southern Brazil is the Lagoa do Peixe National Park, between the cities of Mostardas and Tavares, in Rio Grande do Sul state. This area is a natural reserve implemented to conserve both coastal biodiversity and the many species of migratory birds that use the area in contranuptial periods. Although the flamingo is well known in the region, there is a lack, in scientific literature, of information about the population of flamingos living inside the park. In this paper, we comment on the current population and conservation status of Chilean flamingos in the Lagoa do Peixe National Park, bringing attention to the necessities to protect the park from political pressures and to increase research activity on these birds in this area. Resumen En América del Sur, el flamenco austral (Phoenicopterus chilensis) se distribuye desde el sur del Ecuador hasta el sur de Argentina, pasando por la costa brasileña. -

The Analysis of Destruction in Flamingo Habitat of Acigol Wetland
THE ANALYSIS OF DESTRUCTION IN FLAMINGO HABITAT OF ACIGOL WETLAND M. Karaman*, Z. D. Uca Avci, I. Papila, E. Ozelkan Istanbul Technical University, Center for Satellite Communications and Remote Sensing, Maslak, Istanbul,Turkey (muhittin, damla, emre, papila)@cscrs.itu.edu.tr Abstract: Industrial developments improve human zoologists. The spatial and temporal analyses of water life whereas it can destroy, pollute and decrease the covered areas in the basin area were distinguished by available feeding and breeding area for the animals. considering the feeding type of flamingos as wading. For Acigol Lake, being a saline shallow wetland, offers a the same time period, salt production activities were suitable feeding and breeding area on the migration compared and their effects were evaluated. way of flamingos’. Acigol Lake is the main sodium sulfate production area in Turkey. Sodium sulfate production is carried out by industrial firms and high amounts of water are pumped from lake to salt pans. In this study, spatial and temporal variation of flamingos’ feeding sites due to water pumping is investigated by using remote sensing. Dry and water covered areas were determined by using modified normalized difference water index (MNDWI). Then, the MNDWI images were classified to ‘feeding by wading’ and ‘feeding by swimming’ areas by using the in-situ depth measurements. The decreases in the lake water level during salt production period, showed the Figure 1. The picture of a dead flamingo in Tuz Lake direct effect of industry on flamingos’ feeding areas. (photo credit: Murat Ataol ) (Balkız et al.,2009) Figure 2 . The artemia salina deaths in salt pans Keywords: flamingo, Lake Acigol, MNDWI, salt lake, (Karaman, 2011) salt pans, remote sensing 2. -

Wwtworldwide
In each list below, only one bird has talons to catch its prey. Amazing Adaptations - Answers CIRCLE the bird with talons. Quiz for children aged 5-7 years 9 MALLARD DUCK MUTE SWAN OSPREY 10 MARSH HARRIER REED WARBLER CANADA GOOSE Round 1: Picture round The following birds were all featured in this week’s Amazing Adaptation Cards. Q Can you name them? Round 3: Which is the longest? In this week’s session you looked at birds that have long legs to keep their body above the water. In each list below, CIRCLE the bird that has the longest legs. If you get stuck, use your Amazing Adaptations Cards to help you. 11 REED WARBLER GREY HERON KINGFISHER © Tony Sutton© Tony @ flickr 12 LITTLE EGRET DIPPER MALLARD DUCK 1 (Mute) swan 2 (Grey) heron 3 Avocet 13 DIPPER AVOCET REED WARBLER In this week’s session you looked at birds that had long necks to reach food below the water. In each list below, CIRCLE the bird that has the longest neck. 14 MUTE SWAN OSPREY MALLARD DUCK © ianpreston @ flickr 4 (Mallard) duck 5 Dipper 15 MARSH HARRIER GREY HERON KINGFISHER Round 2: Odd one out round In this week’s session you looked at different types of feet suited to wetlands. In each list below, only one bird has webbed feet. Circle the bird with webbed feet. If you get stuck, use your Amazing Adaptations Cards to help you. Q Can you CIRCLE the odd one out in each list below? 6 REED WARBLER MUTE SWAN OSPREY The otter is a mammal and the other two are amphibians. -

Sparrow Swap: Testing Management Strategies for House Sparrows and Exploring the Use of Their Eggshells for Monitoring Heavy Metal Pollution
ABSTRACT HARTLEY, SUZANNE MARIE. Sparrow Swap: Testing Management Strategies for House Sparrows and Exploring the Use of their Eggshells for Monitoring Heavy Metal Pollution. (Under the direction of Dr. Caren Cooper). Human movement across the globe, particularly through colonialism throughout the last 500 years, has led to the introduction of species into novel environments where they threaten the biodiversity and ecosystem functioning of those novel environments. In the Anthropocene where other threats such as climate change, pollution, and habitat destruction already occur, invasive species are just one more threat facing ecosystems. But what if we can find a way to use an invasive species to help monitor those other threats while at the same time managing them? In the following thesis I explore the strategies by which volunteers manage House Sparrows to minimize their negative impact as an invasive species, but also the potential to use their eggs as indicators of heavy metals in the environment. House Sparrows compete with native birds for nesting spaces. They are also commensal with humans, utilizing buildings as nesting spaces and split grains and forgotten French fries as food sources. In order to 1) find effective management strategies for House Sparrows and 2) evaluate their use as indicators of environmental contaminants, a citizen science project Sparrow Swap was created. Sparrow Swaps takes advantage of the ubiquity of House Sparrows and the expertise of volunteer nestbox monitors to gather data about House Sparrow nesting behaviors and eggs across the United States. In Chapter 1, I address the first research goal of Sparrow Swap by comparing the outcomes of two different strategies by which volunteers manage House Sparrows. -

A Winter Roost of Grey Herons T
A winter roost of Grey Herons T. R. Birkhead INTRODUCTION Gatherings of Grey Herons Ardea cinerea at traditional standing- grounds during the breeding season have been noted by several authors (Beetham 1910, Huxley 1924, Lowe 1954, Milstein et al. 1970), and the behaviour of the birds at these standing-grounds has been investigated (Birkhead 1973). Gatherings of herons outside the breeding season seem to have attracted less attention (see pages 162-163 in Huxley 1924, and Southern 1939), but this may be because they occur less frequently at those times. I consider, how ever, that groups of herons not associated with heronries are less conspicuous and are easily overlooked. The present paper describes observations at a winter day-time communal roost in Yorkshire. Data collected are compared with the results of similar work carried out during the breeding season (Birk head 1973). The roost has been in existence since at least 1966 (personal observation) and probably for considerably longer; B. King (verbally) has recorded a field in Somerset being used by Grey Herons as a roost site for 35 years. ROOST SITE AND OBSERVATIONS The roost site is situated in an area of low-lying arable land in north-west Yorkshire. From September to March, usually each day, herons congregated on the same field, though occasionally either of two adjacent fields was used. The field in which the herons most Fig. 1. Map of the study area, showing the main field used for roosting and the surrounding vegetation Cocksfoot -J Oo Trees 150 j" Observation point 147 148 Winter roost of Grey Herons frequently gathered is 2.63 ha (6.5 acres) in area and slopes gently downwards to the north-west. -

BOURGAS LAST.Pmd
WATERBIRDS OF BOURGAS WETLANDS RESULTS AND EVALUATION OF THE MONTHLY WATERBIRD MONITORING 1996 - 2002 Milko Dimitrov, Tanyo Michev, Lyubomir Profirov, Konstantin Nyagolov 1 WATERBIRDS OF BOURGAS WETLANDS 2 Milko Dimitrov, Tanyo Michev, Lyubomir Profirov, Konstantin Nyagolov WATERBIRDS OF BOURGAS WETLANDS RESULTS AND EVALUATION OF THE MONTHLY WATERBIRD MONITORING 1996 - 2002 Drawings Assen Ignatov Cover design Ivan Yanchev Editor of English Translation Vladimir Pomakov PENSOFT PUBLISHERS BULGARIAN BIODIVERSITY FOUNDATION BOURGAS WETLANDS PUBLICATION SERIES No 6 2005 3 WATERBIRDS OF BOURGAS WETLANDS WATERBIRDS OF BOURGAS WETLANDS Results and Evaluation of the Monthly Waterbird Monitoring 1996 - 2002 Milko Dimitrov, Tanyo Michev, Lyubomir Profirov, Konstantin Nyagolov We dedicate this book to Alexandur Prostov (1926 – 2000), born in Bourgas, who was one of the first to draw the attention of the Bulgarian public to the exceptional richness and diversity of the wildlife and especially the birds of Bourgas Lakes First published 2005 Pensoft Publishers ISBN 954-642-236-3 Pensoft Series Faunistica No 44 ISSN 1312-0174 Bulgarian Biodiversity Foundation ISBN 954-9959-33-3 This book was published with the financial support of the Swiss Agency for Development and Co-operation We recommend this book to be cited in the following way: Dimitrov, M., T. Michev, L. Profirov, K. Nyagolov. 2005. Waterbirds of Bourgas Wetlands. Results and Evaluation of the Monthly Waterbird Monitoring 1996-2002. Bulgarian Biodiversity Foundation and Pensoft Publishers, Sofia–Moscow, 160 pp. On the back cover: Sattelite photo of the region of Bourgas Bay (Original Data © ESA; 1992; Distributed by Eurimage) © PENSOFT Publishers All rights reserved. No part of this publication may be reproduced, stored in a retrieval system or transmitted in any form by any means, electronic, mechanical, photo copying, recording or otherwise, without the prior written permission of the copyright owner.