Transition from Gneiss to Migmatite and the Relationship of Leucosome to Peraluminous Granodiorite in the Cooma Complex, SE Australia R.H
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

JSHESS Early Online View
DOI: 10.22499/3.6901.003 JSHESS early online view This article has been accepted for publication in the Journal of Southern Hemisphere Earth Systems Science and undergone full peer review. It has not been through the copy-editing, typesetting and pagination, which may lead to differences between this version and the final version. A comparison of weather systems in 1870 and 1956 leading to extreme floods in the Murray Darling Basin Jeff Callaghan Retired Bureau of Meteorology Brisbane Queensland Australia. 10 October 2018. 1 ABSTRACT This research is the extension of a project studying the impact of Nineteenth Century severe weather events in Australia and their relation to similar events during the Twentieth and Twenty First Century. Two floods with the worst known impacts in the Murray Darling Basin are studied. One of these events which occurred during 1956 is relatively well known and Bureau of Meteorology archives contain good rainfall data covering the period. Additionally, information on the weather systems causing this rainfall can be obtained. Rainfall, flood and weather system data for this event are presented here and compared with a devastating event during 1870. Although archived Australian rainfall data is negligible during 1870 and there is no record of weather systems affecting Australia during that year, a realistic history of the floods and weather systems in the Murray Darling Basin during 1870 is created. This follows an extensive search through newspaper archives contained in the National Library of Australia’s web site. Examples are presented showing how the meteorological data in Nineteenth Century newspapers can be used to create weather charts. -

Water Sharing Plan for the Murrumbidgee Unregulated and Alluvial Water Sources Amendment Order 2016 Under The
New South Wales Water Sharing Plan for the Murrumbidgee Unregulated and Alluvial Water Sources Amendment Order 2016 under the Water Management Act 2000 I, Niall Blair, the Minister for Lands and Water, in pursuance of sections 45 (1) (a) and 45A of the Water Management Act 2000, being satisfied it is in the public interest to do so, make the following Order to amend the Water Sharing Plan for the Murrumbidgee Unregulated and Alluvial Water Sources 2012. Dated this 29th day of June 2016. NIALL BLAIR, MLC Minister for Lands and Water Explanatory note This Order is made under sections 45 (1) (a) and 45A of the Water Management Act 2000. The object of this Order is to amend the Water Sharing Plan for the Murrumbidgee Unregulated and Alluvial Water Sources 2012. The concurrence of the Minister for the Environment was obtained prior to the making of this Order as required under section 45 of the Water Management Act 2000. 1 Published LW 1 July 2016 (2016 No 371) Water Sharing Plan for the Murrumbidgee Unregulated and Alluvial Water Sources Amendment Order 2016 Water Sharing Plan for the Murrumbidgee Unregulated and Alluvial Water Sources Amendment Order 2016 under the Water Management Act 2000 1 Name of Order This Order is the Water Sharing Plan for the Murrumbidgee Unregulated and Alluvial Water Sources Amendment Order 2016. 2 Commencement This Order commences on the day on which it is published on the NSW legislation website. 2 Published LW 1 July 2016 (2016 No 371) Water Sharing Plan for the Murrumbidgee Unregulated and Alluvial Water Sources Amendment Order 2016 Schedule 1 Amendment of Water Sharing Plan for the Murrumbidgee Unregulated and Alluvial Water Sources 2012 [1] Clause 4 Application of this Plan Omit clause 4 (1) (a) (xxxviii) and (xxxix). -

EIS 968 Environmental Impact Statement for Proposed Sand, Soil and Gravel Extraction at Bredbo in the Shire of Cooma-Monaro
EIS 968 Environmental impact statement for proposed sand, soil and gravel extraction at Bredbo in the Shire of Cooma-Monaro NSW DEPT PEIApy 1NDUSpp1 IIIIIIiu!IIIIIihIIIIih////I/II//II/ll/II///IIjI ABOi 9636 ENVIRONMENTkL IKPACT STAThMENT for proposed Sand, Soil and Gravel Extraction at Bredbo in the Shire of Cooma-Monaro prepared for Lee Aggregates Pty.Ltd. by D.P.JAMES APRIL 1991 Lee Aggregates Pty.Ltd.. D.P.JAMES & COMPANY P.O.Box 397, P.O.Box 170, WANNIASSA, 2903. KOGARAH, 2217. (062)92.3961. (02)588.2614. I I I I I I I I I I I I I I I I I I C ENVIRONMENTAL IMPACT STATEMENT Prepared by D.P.Jaines on behalf of Lee Aggregates Pty.Ltd., P.O.Box 397, Wanniassa, 2903, A.C.T. This is the second edition of this environmental impact statement and is dated April 1991. The first edition is dated June 1988. Minor spelling and typographical errors have been corrected in the second edition, which has been laser printed. '000d-~; 9 April 1991. D.P.James, ARMIT, AMIQ, AIMM. 5/2 Hardie Street, P0 Box 653, 1 NEUTRAL BAY 2089 1- (02)904 1515. / j I 1 INTRODUCTION 1.1 General 1.2 Summary of Proposed Development I 1.3 Development Objectives I 2. EXISTING ENVIRONMENT 2.1 Zoning 22 Landforin 2.3 Land Use I 2.4 Climate & Flooding 2.5 Air Quality 2.6 Water Quality I 2.6.1 Murrunthidgee River 2.7 Noise 2.8 Flora I 2.9 Fauna 2.10 Traffic 2.11 Economic Aspects 2.12 Social & Cultural Aspects Ii 2.13 Archaeology 2.14 Soil & Water Conservation Matters I 2.15 Extractive Industry I ENVIRONMENTAL IMPACTS & PROTECTION MEASURE 3.1 Land Use 3.2 Climate & Flooding 3.3 -

EIS 1505 Proposed Hard Rock Quarry Development, Part Portion 87
EIS 1505 Proposed hard rock quarry development, Part portion 87, Parish of Gladstone, Shire of Cooma-Monaro statement of environmental effects PROPOSED HARD ROCK QUARRY DEVELOPMENT PART PORTION 87. PARISH OF GLADSTONE SHIRE OF COOMA - MONARO STATEMENT OF ENVIRONMENTAL EFFECTS Prepared on behalf of W. and A. Knowles Lm I.T.S. Engineering Services Ply Ltd August 1999 I.TS. Engineering Services Ply. Ltd Civil Engineering Consultants Suite 4 Monaro Centre 108 Commissioner Street COOMA NSW 2630 P0 Box 698 COOMA NSW 2630 Telephone 02 6452 4699 Fax 02 6452 3064 [email protected] PROPOSED QUARRY DEVELOPMENT AT PART PORTION 87 PARISH OF GLADSTONE ROCK FLAT COOMA August 1999 STATEMENT OF ENVIRONMENTAL EFFECTS INDEX 1. THE PROPOSAL 1.1 General Description 1.2 Consideration of Alternatives 1.3 Description of Existing Environment 1.4 Assessment of Environmental Effects DETAILED DESCRIPTION OF THE PROPOSAL 2.1 Aim 2.2 Property Description 2.3 Staging of Development 2.4 Extraction of Material 2.4.1 Type of Material 2.4.2 Area and Quantity of Excavation 2.4.3 Method of Extraction 2.5 Haulage of Material 2.6 Visual Concept 2.7 Final Form of Quany 2.8 Hours of Operation DESCRIPTION OF EXISTING ENVIRONMENT 3.1 Regional Context 3.2 Topography 3.3 Soils 3.4 Hydrology 3.5 Vegetation 3.6 Goulburn-Bombala Railway 3.7 Land Use 3.8 Zoning 3.9 Planning Proposals 3.10 Water quality 3.11 Users of and Demand for Hard Rock Products 3.12 Sources of Hard Rock Product MI11GATING MEASURES 4.1 Visual Impact 4.2 SoilErosion and Water Quality 4.2.1 Diversion of Flows around -

Tectono-Metamorphic Evolution and Timing of The
TECTONO-METAMORPHIC EVOLUTION AND TIMING OF THE MELTING PROCESSES IN THE SVECOFENNIAN TONALITE-TRONDHJEMITE MIGMATITE BELT: AN EXAMPLE FROM LUOPIOINEN, TAMPERE AREA, SOUTHERN FINLAND H. MOURI, K. KORSMÄN and H. HUHMA MOURI, H., KORSMAN, K. and HUHMA. H. 1999. Tectono-metamorphic evolution and timing of the melting processes in the Svecofennian Tonalite- Trondhjemite Migmatite Belt: An example from Luopioinen, Tampere area, southern Finland. Bulletin of the Geological Society of Finland 71, Part 1, 31- 56. The Svecofennian Orogen is in southern Finland characterized by two major migmatite belts. These are the so-called Granite Migmatite Belt, in which Kfs- rich leucosomes predominate, and the Tonalite-Trondhjemite Migmatite Belt, which is characterized by Kfs-poor leucosomes and borders the former belt in the north. The present paper deals with selected migmatitic rocks from the lat- ter belt. It is aimed to study the temporal and structural relationships of the dif- ferent leucosome generations, and to establish the pressure-temperature-time paths of this belt. The Tonalite-Trondhjemite Migmatite Belt consists mainly of migmatitic rocks with various types of synorogenic granitoids and minor mafic and ultramafic rocks. The mesosome of the migmatites consist of garnet-sillimanite-biotite-pla- gioclase-cordierite-quartz assemblages with rare K-feldspar and late andalusite. The oldest leucosomes are dominated by plagioclase and quartz, and the con- tent of K-feldspar increases in later leucosomes. Microtextural analysis in con- junction with THERMOCALC calculations and geothermometry shows that these rocks were metamorphosed at peak conditions of 700-750°C at 4-5 kbar and aH,0 = 0.4-0.7. -

Functioning and Changes in the Streamflow Generation of Catchments
Ecohydrology in space and time: functioning and changes in the streamflow generation of catchments Ralph Trancoso Bachelor Forest Engineering Masters Tropical Forests Sciences Masters Applied Geosciences A thesis submitted for the degree of Doctor of Philosophy at The University of Queensland in 2016 School of Earth and Environmental Sciences Trancoso, R. (2016) PhD Thesis, The University of Queensland Abstract Surface freshwater yield is a service provided by catchments, which cycle water intake by partitioning precipitation into evapotranspiration and streamflow. Streamflow generation is experiencing changes globally due to climate- and human-induced changes currently taking place in catchments. However, the direct attribution of streamflow changes to specific catchment modification processes is challenging because catchment functioning results from multiple interactions among distinct drivers (i.e., climate, soils, topography and vegetation). These drivers have coevolved until ecohydrological equilibrium is achieved between the water and energy fluxes. Therefore, the coevolution of catchment drivers and their spatial heterogeneity makes their functioning and response to changes unique and poses a challenge to expanding our ecohydrological knowledge. Addressing these problems is crucial to enabling sustainable water resource management and water supply for society and ecosystems. This thesis explores an extensive dataset of catchments situated along a climatic gradient in eastern Australia to understand the spatial and temporal variation -

Field Trip Guide: a Profile from Migmatites to Spodumene Pegmatites (Styria, Austria) 1-29
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Berichte der Geologischen Bundesanstalt Jahr/Year: 2019 Band/Volume: 134 Autor(en)/Author(s): Schuster Ralf, Knoll Tanja, Mali Heinrich, Huet Benjamin, Griesmeier Gerit E. U. Artikel/Article: Field trip guide: A profile from migmatites to spodumene pegmatites (Styria, Austria) 1-29 Field trip guide: A profile from migmatites to spodumene pegmatites (Styria, Austria) 7th April 2019 St. Radegund, Austria RALF SCHUSTER, TANJA KNOLL, HEINRICH MALI, BENJAMIN HUET & GERIT E.U. GRIESMEIER Berichte der Geologischen Bundesanstalt, 134 ISSN 1017-8880 Field trip guide: A profile from migmatites to spodumene pegmatites (Styria, Austria) Ralf Schuster, Tanja Knoll, Heinrich Mali, Benjamin Huet & Gerit E.U. Griesmeier Ralf Schuster, Tanja Knoll, Benjamin Huet & Gerit E.U. Griesmeier: Geologische Bundesanstalt, Neulinggasse 38, 1030 Vienna, Austria. Heinrich Mali: Montanuniversität Leoben, Department Applied Geosciences and Geophysics, Peter-Tunner-Straße 5, 8700 Leoben Recommended citation / Zitiervorschlag Schuster, R., Knoll, T., Mali, H., Huet, B. & Griesmeier, G.E.U. (2019): Field trip guide: A profile from migmatites to spodumene pegmatites (Styria, Austria). – Berichte der Geologischen Bundesanstalt, 134, 29 p., Vienna. Cover design: Monika Brüggemann-Ledolter (Geologische Bundesanstalt). Cover pictures: Spodumene pegmatite at Garrach (Styria, Austria) (upper picture: Christine Hörfarter, lower picture: Ralf Schuster) Wien, November 2019 Alle Rechte für das In- und Ausland vorbehalten. © Geologische Bundesanstalt, Wien Technische Redaktion: Christoph Janda Medieninhaber, Herausgeber und Verleger: Geologische Bundesanstalt, Wien Neulinggasse 38, 1030 Wien www.geologie.ac.at Druck: Riegelnik Ges.m.b.H, Piaristengasse 17–19, 1080 Wien Ziel der „Berichte der Geologischen Bundesanstalt“ ist die Verbreitung wissenschaftlicher Ergebnisse durch die Geologische Bundesanstalt. -

Geochemistry and Genesis of Beryl Crystals in the LCT Pegmatite Type, Ebrahim-Attar Mountain, Western Iran
minerals Article Geochemistry and Genesis of Beryl Crystals in the LCT Pegmatite Type, Ebrahim-Attar Mountain, Western Iran Narges Daneshvar 1 , Hossein Azizi 1,* , Yoshihiro Asahara 2 , Motohiro Tsuboi 3 , Masayo Minami 4 and Yousif O. Mohammad 5 1 Department of Mining Engineering, Faculty of Engineering, University of Kurdistan, Sanandaj 66177-15175, Iran; [email protected] 2 Department of Earth and Environmental Sciences, Graduate School of Environmental Studies, Nagoya University, Nagoya 464-8601, Japan; [email protected] 3 Department of Applied Chemistry for Environment, School of Biological and Environmental Sciences, Kwansei Gakuin University, Sanda 669-1337, Japan; [email protected] 4 Division for Chronological Research, Institute for Space-Earth Environmental Research, Nagoya University, Nagoya 464-8601, Japan; [email protected] 5 Department of Geology, College of Science, Sulaimani University, Sulaimani 46001, Iraq; [email protected] * Correspondence: [email protected]; Tel.: +98-918-872-3794 Abstract: Ebrahim-Attar granitic pegmatite, which is distributed in southwest Ghorveh, western Iran, is strongly peraluminous and contains minor beryl crystals. Pale-green to white beryl grains are crystallized in the rim and central parts of the granite body. The beryl grains are characterized by low contents of alkali oxides (Na2O = 0.24–0.41 wt.%, K2O = 0.05–0.17 wt.%, Li2O = 0.03–0.04 wt.%, Citation: Daneshvar, N.; Azizi, H.; and Cs2O = 0.01–0.03 wt.%) and high contents of Be2O oxide (10.0 to 11.9 wt.%). The low contents Asahara, Y.; Tsuboi, M.; Minami, M.; of alkali elements (oxides), low Na/Li (apfu) ratios (2.94 to 5.75), and variations in iron oxide Mohammad, Y.O. -

Table of Contents
Table of Contents Acknowledgements and more………………………………………………………………………….2 Summary of Results.………………………………………………………………………………………..3 Species Detected…………………………………………………………………………………..4 Species Abundance…………………………………………………………………………….…5 Species Results…………………………………………………………………………………………….….9 Discussion…………………………………………………………………………………………….………18 References………………………………………………………………………………………….………...19 Appendix 1 Frogwatch Volunteers 2014………………………………………………….……..20 Appendix 2 Site Location Details………………………………………………………….………...23 Appendix 3 Monitoring Summary 2015…………………………………………………….……42 . 1 Acknowledgements and more This report was written using the data collected by hundreds of ACT and Region FROGWATCH volunteers during October 2014. We would like to offer heartfelt thanks to them, in addition to past program coordinators, who have all shaped and strengthened the FROGWATCH program to what it is today. The immense contributions made by volunteers of all ages and walks of life and from all over the ACT and surrounding region are greatly appreciated and this study would not be possible without them. A special shout out and many thanks go to Will Osborne and to Murray Evans, both have been instrumental in setting up this project and have never stopped supporting it in various ways!! This report was written by: Anke Maria Hoefer, ACT and Region Frogwatch and Sarah Chaplin, volunteer at ACT and Region, Ginninderra Catchment Group. This report can be cited as: Hoefer, A.M. and Chaplin, S.E . (2017): The 2014 FROGWATCH Census Report. Please refer to our online resource page to find out more about the Frogwatch program, its objectives, and the materials and methods used for data collection (http://www.ginninderralandcare.org.au/frogwatch/) 2 Summary of Results: Approximately 144 volunteers monitored 167 sites across the ACT Region during October where 284 surveys were conducted. Of these 284 surveys, 217 were completed during census week (Oct 19-25). -

Migmatites & Their Origin
MIGMATITES & THEIR ORIGIN PAPER- METAMORPHIC PETROLOGY PAPER CODE- MGELCC-7, SEMESTER- 2ND PREPARED BY: Dr. K. Bijendra Pratap Rahi. Guest Assistant Professor Department of Geology Patna University, Patna E mail ID- [email protected] MOBILE NO- 8789608216 • Definition: • Sederholm (1908) introduced the term migmatite to designate certain gneissic rocks “which look like mixed rocks”. • Coarse grained, heterogeneous and megascopically composite rocks, comprising portions of granitic material (quartz + feldspar) called leucosome, and dark coloured portions of the metamorphic rocks containing mafic minerals, e.g., biotite, garnet, sillimanite, cordierite etc., which is called melanosome. • lit-par-lit gneiss, composite gneiss, injection gneiss etc. • Field relations and Occurrence: • Migmatites are associated with metamorphic rocks of the highest temperature part of the amphibolite facies and granulite facies. • “One of the most firmly established facts in metamorphic geology is the close association in field of highest grade metamorphic rocks and migmatites” Read (1957) • This worldwide observation suggests that the spatial association of these rocks is due to processes occurring at similar temperature and pressure conditions in the deeper part of the crust and giving rise to high-grade metamorphic rocks and migmatites. • The origin of migmatites in deep seated parts of orogenic belts must be considered as directly connected with high-grade metamorphism. Origin of migmatites in light of experimental work: • Shales and greywackes are common sediments deposited in a geosyncline. • The metamorphism of these sediments was experimentally investigated by Winkler and his coworkers. • The experimental anatexis of quartz-albite-K- feldspar (granite) system was investigated by Tuttle and Bowen(1958) and their data are plotted in Fig.1. -
Answers to Questions on Notice
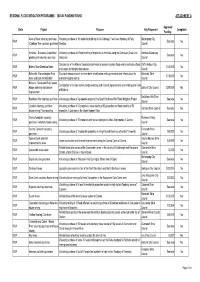
REGIONAL FLOOD MITIGATION PROGRAMME - 2003-04 FUNDING ROUND ATTACHMENT A Approved State Project Purpose Key Proponent Completed Funding Anama Street voluntary purchase Voluntary purchase of 16 residential buildings in the Cabbage Tree Creek floodway of Fairy Wollongong City NSW See note No (Cabbage Tree voluntary purchase) Meadow. Council Armidale - Dumaresq Creek flood Voluntary purchase and flood proofing of properties in Armidale along the Dumaresq Creek and Armidale Dumaresq NSW See note No proofing and voluntary purchase tributaries. Council Construciton of the Bakers Road detention basin to temporarily store flood waters and reduce flood Coffs Harbour City NSW Bakers Road Detention Basin $140,000 No discharges and heights downstream. Council Balranald - Murrumbidgee River Structural measures such as levee bank rehabilitation and augmentation and infrastructure for Balranald Shire NSW $110,000 No levee and bank rehabilitation overland flooding control. Council Bathurst - Macquarie River levees, Construction of multiple levees, bridge widening and channel improvements on the Macquarie River NSW bridge widening and channel Bathurst City Council $300,000 No at Bathurst. improvement Baulkham Hills Shire NSW Baulkham Hills voluntary purchase Voluntary purchase of 6 properties as part of the South Baulkham Hills Flood Mitigation Project. See note No Council Camden voluntary purchase / Voluntary purchase of 43 properties, house raising of 62 properties and flood proofing of 56 NSW Camden Shire Council See note No house raising / flood proofing properties in Camden on the Upper Nepean River. Casino floodplain voluntary Richmond Valley NSW Voluntary purchase of 12 properties and house raising of a further 26 properties in Casino. See note No purchase / voluntary house raising Council Central Tamworth voluntary Tamworth Shire NSW Voluntary purchase of 2 residential properties in a high hazard flood way of central Tamworth. -

Geochemical Characterization of Migmatites in Funtua Sheet 78 North-East, Scale 1:50,000 North Western Nigeria
International Journal of Innovative Research in Education, Technology & Social Strategies p-ISSN: 2465-7298 | e-ISSN: 2467-8163 IJIRETSS Volume 7, Number 1 February, 2020 Geochemical Characterization of Migmatites in Funtua Sheet 78 North-East, Scale 1:50,000 North Western Nigeria 1Kankara, I. A. & 2Galadanchi, K. M. 1Department of Geology, Federal University Dutsin-Ma 2Department of Chemistry, Umaru Musa Yar'adua University, Katsina A b s t r a c t n this present study detailed chemical analyses for major and trace elements were done on a total of 20 samples of the rocks investigated using the AAS, IXRF and FP (Flame Photometry) The study area is shown to extend into the north western areas of northern Nigeria and even beyond. The age of D1 and D2 deformations and metamorphisms in the FTNE is therefore shown to be Pan- African. At least two geotectonic episodes or intense phases of ductile deformation were identified in the field to have affected the rocks. The rocks have been strongly foliated with crystals showing strong preferred orientations. Fresh and partly weathered rocks exhibited on the extent and effects of wall-rock alterations that must have accompanied the Pan-African orogeny. The harmonized Analytical results showed that the rocks are saturated with respect to silica, and contain moderate to elevated concentrations of Al2O3, comparable to most pelitic rocks. There appears to be no consistency in the relative concentration of Alkalis in the rocks, and this possibly indicates a mixed nature of the source materials. Both MgO and CaO compositions portray sedimentary character of the protoliths of the rocks.