Observations on Stem Surface Anatomy In
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Alberta Wetland Classification System – June 1, 2015
Alberta Wetland Classification System June 1, 2015 ISBN 978-1-4601-2257-0 (Print) ISBN: 978-1-4601-2258-7 (PDF) Title: Alberta Wetland Classification System Guide Number: ESRD, Water Conservation, 2015, No. 3 Program Name: Water Policy Branch Effective Date: June 1, 2015 This document was updated on: April 13, 2015 Citation: Alberta Environment and Sustainable Resource Development (ESRD). 2015. Alberta Wetland Classification System. Water Policy Branch, Policy and Planning Division, Edmonton, AB. Any comments, questions, or suggestions regarding the content of this document may be directed to: Water Policy Branch Alberta Environment and Sustainable Resource Development 7th Floor, Oxbridge Place 9820 – 106th Street Edmonton, Alberta T5K 2J6 Phone: 780-644-4959 Email: [email protected] Additional copies of this document may be obtained by contacting: Alberta Environment and Sustainable Resource Development Information Centre Main Floor, Great West Life Building 9920 108 Street Edmonton Alberta Canada T5K 2M4 Call Toll Free Alberta: 310-ESRD (3773) Toll Free: 1-877-944-0313 Fax: 780-427-4407 Email: [email protected] Website: http://esrd.alberta.ca Alberta Wetland Classification System Contributors: Matthew Wilson Environment and Sustainable Resource Development Thorsten Hebben Environment and Sustainable Resource Development Danielle Cobbaert Alberta Energy Regulator Linda Halsey Stantec Linda Kershaw Arctic and Alpine Environmental Consulting Nick Decarlo Stantec Environment and Sustainable Resource Development would also -

Clonal Structure in the Moss, Climacium Americanum Brid
Heredity 64 (1990) 233—238 The Genetical Society of Great Britain Received 25August1989 Clonal structure in the moss, Climacium americanum Brid. Thomas R. Meagher* and * Departmentof Biological Sciences, Rutgers University, P.O. Box 1059, Jonathan Shawt Piscataway, NJ 08855-1059. 1 Department of Biology, Ithaca College, Ithaca, NY 14850. The present study focussed on the moss species, Climacium americanum Brid., which typically grows in the form of mats or clumps of compactly spaced gametophores. In order to determine the clonal structure of these clumps, the distribution of genetic variation was analysed by an electrophoretic survey. Ten individual gametophores from each of ten clumps were sampled in two locations in the Piedmont of North Carolina. These samples were assayed for allelic variation at six electrophoretically distinguishable loci. For each clump, an explicit probability of a monomorphic versus a polymorphic sample was estimated based on allele frequencies in our overall sample. Although allelic variation was detected within clumps at both localities, our statistical results showed that the level of polymorphism within clumps was less than would be expected if genetic variation were distributed at random within the overall population. INTRODUCTION approximately 50 per cent of moss species that are hermaphroditic, self-fertilization of bisexual Ithas been recognized that in many taxa asexual gametophytes results in completely homozygous propagation as the predominant means of repro- sporophytes. Dispersal of such genetically -

About the Book the Format Acknowledgments
About the Book For more than ten years I have been working on a book on bryophyte ecology and was joined by Heinjo During, who has been very helpful in critiquing multiple versions of the chapters. But as the book progressed, the field of bryophyte ecology progressed faster. No chapter ever seemed to stay finished, hence the decision to publish online. Furthermore, rather than being a textbook, it is evolving into an encyclopedia that would be at least three volumes. Having reached the age when I could retire whenever I wanted to, I no longer needed be so concerned with the publish or perish paradigm. In keeping with the sharing nature of bryologists, and the need to educate the non-bryologists about the nature and role of bryophytes in the ecosystem, it seemed my personal goals could best be accomplished by publishing online. This has several advantages for me. I can choose the format I want, I can include lots of color images, and I can post chapters or parts of chapters as I complete them and update later if I find it important. Throughout the book I have posed questions. I have even attempt to offer hypotheses for many of these. It is my hope that these questions and hypotheses will inspire students of all ages to attempt to answer these. Some are simple and could even be done by elementary school children. Others are suitable for undergraduate projects. And some will take lifelong work or a large team of researchers around the world. Have fun with them! The Format The decision to publish Bryophyte Ecology as an ebook occurred after I had a publisher, and I am sure I have not thought of all the complexities of publishing as I complete things, rather than in the order of the planned organization. -

Household and Personal Uses
Glime, J. M. 2017. Household and Personal Uses. Chapt. 1-1. In: Glime, J. M. Bryophyte Ecology. Volume 5. Uses. Ebook sponsored 1-1-1 by Michigan Technological University and the International Association of Bryologists. Last updated 5 October 2017 and available at <http://digitalcommons.mtu.edu/bryophyte-ecology/>. CHAPTER 1 HOUSEHOLD AND PERSONAL USES TABLE OF CONTENTS Household Uses...................................................................................................................................................1-1-2 Furnishings...................................................................................................................................................1-1-4 Padding and Absorption...............................................................................................................................1-1-5 Mattresses.............................................................................................................................................1-1-6 Shower Mat...........................................................................................................................................1-1-7 Urinal Absorption.................................................................................................................................1-1-8 Cleaning.......................................................................................................................................................1-1-8 Brushes and Brooms.............................................................................................................................1-1-8 -

<I>Sphagnum</I> Peat Mosses
ORIGINAL ARTICLE doi:10.1111/evo.12547 Evolution of niche preference in Sphagnum peat mosses Matthew G. Johnson,1,2,3 Gustaf Granath,4,5,6 Teemu Tahvanainen, 7 Remy Pouliot,8 Hans K. Stenøien,9 Line Rochefort,8 Hakan˚ Rydin,4 and A. Jonathan Shaw1 1Department of Biology, Duke University, Durham, North Carolina 27708 2Current Address: Chicago Botanic Garden, 1000 Lake Cook Road Glencoe, Illinois 60022 3E-mail: [email protected] 4Department of Plant Ecology and Evolution, Evolutionary Biology Centre, Uppsala University, Norbyvagen¨ 18D, SE-752 36, Uppsala, Sweden 5School of Geography and Earth Sciences, McMaster University, Hamilton, Ontario, Canada 6Department of Aquatic Sciences and Assessment, Swedish University of Agricultural Sciences, SE-750 07, Uppsala, Sweden 7Department of Biology, University of Eastern Finland, P.O. Box 111, 80101, Joensuu, Finland 8Department of Plant Sciences and Northern Research Center (CEN), Laval University Quebec, Canada 9Department of Natural History, Norwegian University of Science and Technology University Museum, Trondheim, Norway Received March 26, 2014 Accepted September 23, 2014 Peat mosses (Sphagnum)areecosystemengineers—speciesinborealpeatlandssimultaneouslycreateandinhabitnarrowhabitat preferences along two microhabitat gradients: an ionic gradient and a hydrological hummock–hollow gradient. In this article, we demonstrate the connections between microhabitat preference and phylogeny in Sphagnum.Usingadatasetof39speciesof Sphagnum,withan18-locusDNAalignmentandanecologicaldatasetencompassingthreelargepublishedstudies,wetested -

Climacium Dendroides from Cofre De Perote, a High-Elevation Tropical Montane Site in Veracruz, Mexico
The Bryologist 107(3), pp. 368 372 Copyright q 2004 by the American Bryological and Lichenological Society, Inc. Climacium dendroides from Cofre de Perote, A High-Elevation Tropical Montane Site in Veracruz, Mexico ANGELA E. NEWTON Department of Botany, Natural History Museum, London SW7 5BD, United Kingdom; e-mail: a.newton@nhm. ac.uk EFRAIÂN DE LUNA Instituto de EcologõÂa, A.C., Ap. Postal 63, 91000 Xalapa, Veracruz, MeÂxico; e-mail: [email protected] Abstract. Climacium dendroides (Hedw.) F. Weber & D. Mohr is reported new to Mexico and in the ®rst known locality in the tropics, at the elevation of 3,000 m, on the ancient shield volcano of Cofre de Perote. Variation in branching of the costa is assessed in this plant and in material of C. dendroides worldwide. Additional taxa of interest from this locality on Cofre de Perote include Aulacomnium palustre and Polytrichum commune (both new to the state of Veracruz). Keywords. `Boreal' mosses, Climacium, distribution, Mexico, montane tropics. The ancient shield volcano of Cofre de Perote is 20 m across, and consisted of a series of small located in the Neovolcanic Axis of Mexico, at swamps connected by the small stream. Climacium 198329 N, 978309 W, and reaches an elevation of was growing on low hummocks to one side of the 4,200 m. The upper regions of the mountain have stream, and the plants were deeply embedded in the extensive Pinus-Abies forests with a rich ¯ora of surrounding mosses and other plants. Additional mosses, ferns, and cycads, that has been the focus moss species in the immediate area included Po- of much botanical study in the past, including the lytrichum commune Hedw. -

New York Natural Heritage Program
Kohler Environmental Center Ecological Community Classification and Mapping New York Natural Heritage Program The NY Natural Heritage Program is a partnership between the NYS Department of Environmental Conservation (NYSDEC) and the State University of New York College of Environmental Science and Forestry. New York Natural Heritage Program 625 Broadway, 5th Floor Albany, NY 12233-4757 (518) 402-8935 Fax (518) 402-8925 www.nynhp.org Kohler Environmental Center Ecological Community Classification and Mapping Gregory J. Edinger A report prepared by the New York Natural Heritage Program 625 Broadway, 5th Floor Albany, NY 12233-4757 www.nynhp.org for Choate Rosemary Hall 333 Christian Street. Wallingford, CT 06492 January 2014 Please cite this report as follows: Edinger, G.J. 2014. Kohler Environmental Center Ecological Community Classification and Mapping. New York Natural Heritage Program, Albany, NY. Cover photos: top left: shallow emergent marsh (CEGL006446) at point A06; top right: red maple-hardwood swamp (CEGL006406) at point C03; bottom: coastal oak-beech forest (CEGL006377) at point C02. New York Natural Heritage Program Table of Contents INTRODUCTION ........................................................................................................................................... 3 Purpose and Study Area Overview ............................................................................................................. 3 Study Area Physical Setting and Environment ......................................................................................... -

A Molecular Phylogeny of Hypnales (Bryophyta) Inferred from ITS2 Sequence-Structure Data Benjamin Merget, Matthias Wolf*
Merget and Wolf BMC Research Notes 2010, 3:320 http://www.biomedcentral.com/1756-0500/3/320 SHORT REPORT Open Access A molecular phylogeny of Hypnales (Bryophyta) inferred from ITS2 sequence-structure data Benjamin Merget, Matthias Wolf* Abstract Background: Hypnales comprise over 50% of all pleurocarpous mosses. They provide a young radiation complicating phylogenetic analyses. To resolve the hypnalean phylogeny, it is necessary to use a phylogenetic marker providing highly variable features to resolve species on the one hand and conserved features enabling a backbone analysis on the other. Therefore we used highly variable internal transcribed spacer 2 (ITS2) sequences and conserved secondary structures, as deposited with the ITS2 Database, simultaneously. Findings: We built an accurate and in parts robustly resolved large scale phylogeny for 1,634 currently available hypnalean ITS2 sequence-structure pairs. Conclusions: Profile Neighbor-Joining revealed a possible hypnalean backbone, indicating that most of the hypnalean taxa classified as different moss families are polyphyletic assemblages awaiting taxonomic changes. Background encompassing a total of 1,634 species in order to test Pleurocarpous mosses, which are mainly found in tropi- the hypothesis that the ITS2 sequence-structure can be cal forests, account for more than 50% of all moss spe- used to determine the phylogeny of Hypnales and to cies [1,2]. Brotherus in 1925 used morphological resolve especially its phylogenetic backbone. A rapid characters to partition the pleurocarpous into three radiation in the early history of pleurocarpous mosses orders. These were Leucodontales (= Isobryales), Hoo- has resulted in low molecular diversity generally, but keriales and Hypnobryales (= Hypnales) [3]. Later mole- particularly in the order Hypnales [5,7]. -

Field Guide to the Moss Genera in New Jersey by Keith Bowman
Field Guide to the Moss Genera in New Jersey With Coefficient of Conservation and Indicator Status Keith Bowman, PhD 10/20/2017 Acknowledgements There are many individuals that have been essential to this project. Dr. Eric Karlin compiled the initial annotated list of New Jersey moss taxa. Second, I would like to recognize the contributions of the many northeastern bryologists that aided in the development of the initial coefficient of conservation values included in this guide including Dr. Richard Andrus, Dr. Barbara Andreas, Dr. Terry O’Brien, Dr. Scott Schuette, and Dr. Sean Robinson. I would also like to acknowledge the valuable photographic contributions from Kathleen S. Walz, Dr. Robert Klips, and Dr. Michael Lüth. Funding for this project was provided by the United States Environmental Protection Agency, Region 2, State Wetlands Protection Development Grant, Section 104(B)(3); CFDA No. 66.461, CD97225809. Recommended Citation: Bowman, Keith. 2017. Field Guide to the Moss Genera in New Jersey With Coefficient of Conservation and Indicator Status. New Jersey Department of Environmental Protection, New Jersey Forest Service, Office of Natural Lands Management, Trenton, NJ, 08625. Submitted to United States Environmental Protection Agency, Region 2, State Wetlands Protection Development Grant, Section 104(B)(3); CFDA No. 66.461, CD97225809. i Table of Contents Introduction .................................................................................................................................................. 1 Descriptions -

Water Relations: Conducting Structures
Glime, J. M. 2017. Water Relations: Conducting Structures. Chapt. 7-1. In: Glime, J. M. Bryophyte Ecology. Volume 1. Physiological 7-1-1 Ecology. Ebook sponsored by Michigan Technological University and the International Association of Bryologists. Last updated 7 March 2017 and available at <http://digitalcommons.mtu.edu/bryophyte-ecology/>. CHAPTER 7-1 WATER RELATIONS: CONDUCTING STRUCTURES TABLE OF CONTENTS Movement to Land .............................................................................................................................................. 7-1-2 Bryophytes as Sponges ....................................................................................................................................... 7-1-2 Conducting Structure .......................................................................................................................................... 7-1-4 Leptomes and Hydromes............................................................................................................................. 7-1-8 Hydroids............................................................................................................................................. 7-1-12 Leptoids.............................................................................................................................................. 7-1-14 Rhizome..................................................................................................................................................... 7-1-15 Leaves....................................................................................................................................................... -

A Baseline Study of Alpine Snowbed and Rill Communities on Mount Washington, Nh Author(S): Robert S
A Baseline Study of Alpine Snowbed and Rill Communities On Mount Washington, Nh Author(s): Robert S. Capers Nancy G. Slack Source: Rhodora, 118(976):345-381. Published By: The New England Botanical Club, Inc. DOI: http://dx.doi.org/10.3119/16-07 URL: http://www.bioone.org/doi/full/10.3119/16-07 BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use. Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. RHODORA, Vol. 118, No. 976, pp. 345–381, 2016 Ó Copyright 2016 by the New England Botanical Club doi: 10.3119/16-07; first published on-line February 6, 2017. A BASELINE STUDY OF ALPINE SNOWBED AND RILL COMMUNITIES ON MOUNT WASHINGTON, NH ROBERT S. CAPERS Department of Ecology and Evolutionary Biology, University of Connecticut, Storrs, CT 06269 e-mail: [email protected] NANCY G. SLACK Biology Department, The Sage Colleges, 65 1st St., Troy, NY 12180 ABSTRACT. -
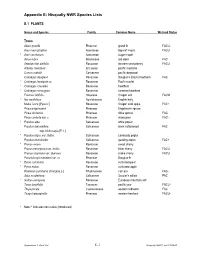
Appendix E Nisqually Species List
Appendix E: Nisqually NWR Species Lists E.1 PLANTS Genus and Species Family Common Name Wetland Status Trees Abies grandis Pinaceae grand fir FACU- Acer macrophyllum Aceraceae big-leaf maple FACU * Acer saccharum Aceraceae sugar maple Alnus rubra Betulaceae red alder FAC Amelanchier alnifolia Rosaceae western serviceberry FACU Arbutus menziesii Ericaceae pacific madrone Cornus nuttallii Cornaceae pacific dogwood Crataegus douglasii Rosaceae Douglas’s (black) hawthorn FAC * Crataegus laevigata cv. Rosaceae Paul's scarlet * Crataegus x lavallei Rosaceae hawthorn * Crataegus monogyna Rosaceae common hawthorn Fraxinus latifolia Oleaceae Oregon ash FACW * Ilex aquifolium Aquifoliaceae English holly Malus fusca [Pyrus f.] Rosaceae Oregon crab apple FAC+ Picea engelmannii Pinaceae Engelmann spruce Picea sitchensis Pinaceae Sitka spruce FAC Pinus contorta var. c. Pinaceae shore pine FAC- * Populus alba Salicaceae white poplar Populus balsamifera Salicaceae black cottonwood FAC ssp. trichocarpa [P. t. ] * Populus nigra var. italica Salicaceae Lombardy poplar Populus tremuloides Salicaceae quaking aspen FAC+ * Prunus avium Rosaceae sweet cherry Prunus emarginata var. mollis Rosaceae bitter cherry FACU Prunus virginiana var. demissa Rosaceae choke cherry FACU Pseudotsuga menziesii var. m. Pinaceae Douglas-fir * Pyrus communis Rosaceae cultivated pear * Pyrus malus Rosaceae cultivated apple Rhamnus purshiana [Frangula p.] Rhamnaceae cascara FAC- Salix scouleriana Salicaceae Scouler’s willow FAC * Sorbus aucuparia Rosaceae European mountain ash Taxus brevifolia Taxaceae pacific yew FACU- Thuja plicata Cupressaceae western redcedar FAC Tsuga heterophylla Pinaceae western hemlock FACU- * Note: * indicates non-native (introduced) Appendix E.1: Plant List E-1 Nisqually NWR Final CCP/EIS Genus and Species Family Common Name Wetland Status Shrubs, Brambles & Vines Acer circinatum Aceraceae vine maple FACU+ Arctostaphylos uva-ursi var.