Cytoplasmic Inheritance of Plastids in Impatiens Sultanii Hook, F., Petunia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

State of New York City's Plants 2018
STATE OF NEW YORK CITY’S PLANTS 2018 Daniel Atha & Brian Boom © 2018 The New York Botanical Garden All rights reserved ISBN 978-0-89327-955-4 Center for Conservation Strategy The New York Botanical Garden 2900 Southern Boulevard Bronx, NY 10458 All photos NYBG staff Citation: Atha, D. and B. Boom. 2018. State of New York City’s Plants 2018. Center for Conservation Strategy. The New York Botanical Garden, Bronx, NY. 132 pp. STATE OF NEW YORK CITY’S PLANTS 2018 4 EXECUTIVE SUMMARY 6 INTRODUCTION 10 DOCUMENTING THE CITY’S PLANTS 10 The Flora of New York City 11 Rare Species 14 Focus on Specific Area 16 Botanical Spectacle: Summer Snow 18 CITIZEN SCIENCE 20 THREATS TO THE CITY’S PLANTS 24 NEW YORK STATE PROHIBITED AND REGULATED INVASIVE SPECIES FOUND IN NEW YORK CITY 26 LOOKING AHEAD 27 CONTRIBUTORS AND ACKNOWLEGMENTS 30 LITERATURE CITED 31 APPENDIX Checklist of the Spontaneous Vascular Plants of New York City 32 Ferns and Fern Allies 35 Gymnosperms 36 Nymphaeales and Magnoliids 37 Monocots 67 Dicots 3 EXECUTIVE SUMMARY This report, State of New York City’s Plants 2018, is the first rankings of rare, threatened, endangered, and extinct species of what is envisioned by the Center for Conservation Strategy known from New York City, and based on this compilation of The New York Botanical Garden as annual updates thirteen percent of the City’s flora is imperiled or extinct in New summarizing the status of the spontaneous plant species of the York City. five boroughs of New York City. This year’s report deals with the City’s vascular plants (ferns and fern allies, gymnosperms, We have begun the process of assessing conservation status and flowering plants), but in the future it is planned to phase in at the local level for all species. -

Mirabilis Jalapa: a Review of Ethno and Pharmacological Activities
Advancement in Medicinal Plant Research Vol. 9(1), pp. 1-10, January 2021 ISSN: 2354-2152 Review Mirabilis jalapa: A review of ethno and pharmacological activities Farjana Islam Liya, Mt. Farzana Yasmin, Nargis Sultana Chowdhury*, Tasnia Khasru Charu and Ismat Benta Fatema Department of Pharmacy, Manarat International University, Ashulia Model Town, Khagan, Ashulia, Dhaka, Bangladesh. Accepted 4 January, 2021 ABSTRACT Plants have been used for thousands of years to treat, prevent, and control a variety of diseases throughout the world. The initial benefits of using plant-derived medicine are that they are relatively safer than artificial alternatives. Mirabilis jalapa Linn. (Nyctaginaceae) is one of the plants that used for health care and medicinal purposes for several thousands of years. It is a perennial bushy herb promulgate by flowers or leaves, a native of America and commonly known as ‘four-o-clock’. It has traditionally been used in the treatment of gastrointestinal disorders, muscle pain, abdominal colic, and diarrhea in ancient Mexico, Japan, China and Brazil. The literature review revealed that M. jalapa is widely used as anti-oxidant, anti- inflammatory, anti-microbial, anti-diabetic, cytotoxic, antinociceptive, and several other’s medicines. In aerial parts of the M. jalapa, triterpene and flavonoids are found. Flowers mostly contain anthocyanins and flavonoids. Carbohydrate, resin and alkaloids are found in roots. Tricosan-12-one, n-hexacosanal, β- sitosterol, and tetracosanoic were isolated from the leaves of the M. jalapa. Seeds contain β-sitosterol, β- amyrin and β-sitosterol-D-glucoside. The presence of various bioactive compounds validates the whole plant for different medicinal practitioners. -
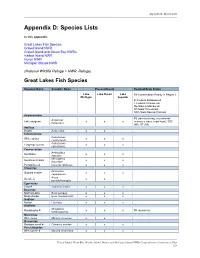
Species Lists
Appendix D: Species Lists Appendix D: Species Lists In this appendix: Great Lakes Fish Species Gravel Island NWR Gravel Island and Green Bay NWRs Harbor Island NWR Huron NWR Michigan Islands NWR (National Wildlife Refuge = NWR, Refuge) Great Lakes Fish Species Common Name Scientific Name Present/Absent Regional/State Status Lake Lake Huron Lake R3-Conservation Priority in Region 3 Michigan Superior E- Federal Endangered T-Federal Threatened SE-State Endangered ST-State Threatened SSC-State Special Concern Acipenseridae R3 (rare/declining, recreational/ Acipenser Lake sturgeon x x x economic value, tribal trust), SSC fulvescens (WI), ST (MI) Amiidae Bowfin Amia calva x x Catostomidae Catostomus White sucker x x x commersoni Catostomus Longnose sucker x x x catostomus Centrarchidae Ambloplites Rockbass x x x rupestris Micropterus Smallmouth bass x x x dolomieui Pumpkinseed Lepomis gibbosus x x x Clupeidae Dorosoma Gizzard shad # x x x cepedianum Alosa Alewife # x x pseudoharengus Cyprinidae Carp # Cyprinus Carpio x x x Esocidae Northern pike Esox Lucieus x x x Muskellunge Esox masquinongy x x x Gadidae Burbot Lota lota x x x Gobiidae Neogobius Round goby # x x x R3 (nuisance) melanostomus Moronidae White bass Morone chrysops x x Osmeridae Rainbow smelt # Osmerus mordax x x x Percichthyidae White perch # Morone americana x x x Gravel Island, Green Bay, Harbor Island, Huron, and Michigan Islands NWRs/Comprehensive Conservation Plan 221 Appendix D: Species Lists Common Name Scientific Name Present/Absent Regional/State Status Percidae R3 (rare/declining, -

ANNUALS for UTAH GARDENS Teresa A
ANNUALS FOR UTAH GARDENS Teresa A. Cerny Ornamental Horticulture Specialist Debbie Amundsen Davis County Horticulture Extension Agent Loralie Cox Cache County Horticulture Extension Agent September 2003 HG-2003/05 Annuals are plants that come up in the spring, reach maturity, flower, set seeds, then die all in one season. They provide eye-catching color to any flower bed and can be used as borders, fillers, or background plantings. There are several ways to find annual species that fit your landscape needs; referring to the All-American Selection program evaluations (http://www.all-americaselections.org), visiting botanical gardens to observe examples of annuals in the landscape, and looking through commercial seed catalogs are excellent places to find ideas. Most annuals are available in cell packs, flats, or individual pots. When buying plants, choose those that are well established but not pot bound. Tall spindly plants lack vigor and should be avoided. Instead look for plants with dark green foliage that are compact and free of insect and disease problems. These criteria are much more important than the flower number when choosing a plant. An abundance of foliage with few, if any flowers, is desirable. BED PREPARATION Avoid cultivating soil too early in the spring and during conditions that are too wet. Soil conditions can be determined by feeling the soil. If the soil forms a ball in your hand but crumbles easily, it is ideal. Cultivate the flower bed to a depth of 6-10 inches by turning the soil with a spade. Utah soils can always use extra organic matter such as grass clippings, leaves, compost, manure, peat, etc. -

Illinois Exotic Species List
Exotic Species in Illinois Descriptions for these exotic species in Illinois will be added to the Web page as time allows for their development. A name followed by an asterisk (*) indicates that a description for that species can currently be found on the Web site. This list does not currently name all of the exotic species in the state, but it does show many of them. It will be updated regularly with additional information. Microbes viral hemorrhagic septicemia Novirhabdovirus sp. West Nile virus Flavivirus sp. Zika virus Flavivirus sp. Fungi oak wilt Ceratocystis fagacearum chestnut blight Cryphonectria parasitica Dutch elm disease Ophiostoma novo-ulmi and Ophiostoma ulmi late blight Phytophthora infestans white-nose syndrome Pseudogymnoascus destructans butternut canker Sirococcus clavigignenti-juglandacearum Plants okra Abelmoschus esculentus velvet-leaf Abutilon theophrastii Amur maple* Acer ginnala Norway maple Acer platanoides sycamore maple Acer pseudoplatanus common yarrow* Achillea millefolium Japanese chaff flower Achyranthes japonica Russian knapweed Acroptilon repens climbing fumitory Adlumia fungosa jointed goat grass Aegilops cylindrica goutweed Aegopodium podagraria horse chestnut Aesculus hippocastanum fool’s parsley Aethusa cynapium crested wheat grass Agropyron cristatum wheat grass Agropyron desertorum corn cockle Agrostemma githago Rhode Island bent grass Agrostis capillaris tree-of-heaven* Ailanthus altissima slender hairgrass Aira caryophyllaea Geneva bugleweed Ajuga genevensis carpet bugleweed* Ajuga reptans mimosa -

Biological Activities of Trifolium Pratense: a Review
Acta Scientific Pharmaceutical Sciences (ISSN: 2581-5423) Volume 3 Issue 9 September 2019 Review Article Biological Activities of Trifolium Pratense: A Review Atiq-ur-Rehman1,2* 1University College of Pharmacy, University of the Punjab, Lahore, Pakistan 2Faculty of Pharmacy, The University of Lahore, Lahore, Pakistan *Corresponding Author: Atiq-ur-Rehman, Faculty of Pharmacy, The University of Lahore and University College of Pharmacy, University of the Punjab, Lahore Pakistan. Received: July 25, 2019; Published: August 16, 2019 Abstract Trifolium pratense is an important plant of the Legume family. It has drawn the attention of several researchers around the globe. This plant was traditionally used as forage or as soil improver is now seen as the plant containing vast therapeutic activities which include anti-oxidative, anti-cancer, neuroprotective, anti-hyperglycemic, anti-hyperlipidemic, osteoprotective and cardio protective properties. The therapeutic properties are shown in various in vivo, in vitro and ex vivo experiments. The review highlights the Tri- forium pratense basic knowledge its extraction, components and their actions, major activities possessed by plant along with their mechanisms. Trifolium plant is mainmajorly used in menopausal women to reduce the discomfort and menopausal effects such as moderate cancer causing cells. Various strategies were applied and the plant is still under study for further development in its effects. hot flushes and increase in breast density. The plant is also majorly responsible for preventing breast cancer and other apoptosis of Keywords: Trifolium Pratense; Cancer; Trifolium Introduction Family The genus Trifolium comprises of almost 240 species each re- It belongs to the family Fabeaceae leguminosae. markable for its agricultural and therapeutic effects. -

Atoll Research Bulletin No. 503 the Vascular Plants Of
ATOLL RESEARCH BULLETIN NO. 503 THE VASCULAR PLANTS OF MAJURO ATOLL, REPUBLIC OF THE MARSHALL ISLANDS BY NANCY VANDER VELDE ISSUED BY NATIONAL MUSEUM OF NATURAL HISTORY SMITHSONIAN INSTITUTION WASHINGTON, D.C., U.S.A. AUGUST 2003 Uliga Figure 1. Majuro Atoll THE VASCULAR PLANTS OF MAJURO ATOLL, REPUBLIC OF THE MARSHALL ISLANDS ABSTRACT Majuro Atoll has been a center of activity for the Marshall Islands since 1944 and is now the major population center and port of entry for the country. Previous to the accompanying study, no thorough documentation has been made of the vascular plants of Majuro Atoll. There were only reports that were either part of much larger discussions on the entire Micronesian region or the Marshall Islands as a whole, and were of a very limited scope. Previous reports by Fosberg, Sachet & Oliver (1979, 1982, 1987) presented only 115 vascular plants on Majuro Atoll. In this study, 563 vascular plants have been recorded on Majuro. INTRODUCTION The accompanying report presents a complete flora of Majuro Atoll, which has never been done before. It includes a listing of all species, notation as to origin (i.e. indigenous, aboriginal introduction, recent introduction), as well as the original range of each. The major synonyms are also listed. For almost all, English common names are presented. Marshallese names are given, where these were found, and spelled according to the current spelling system, aside from limitations in diacritic markings. A brief notation of location is given for many of the species. The entire list of 563 plants is provided to give the people a means of gaining a better understanding of the nature of the plants of Majuro Atoll. -

Induction and Maintenance of Callus from Leaf Explants of Mirabilis Jalapa L
® Medicinal and Aromatic Plant Science and Biotechnology ©2009 Global Science Books Induction and Maintenance of Callus from Leaf Explants of Mirabilis jalapa L. Anna Pick Kiong Ling1* • Kuok-Yid Tang1 • Jualang Azlan Gansau2 • Sobri Hussein3 1 Department of Bioscience, Faculty of Engineering and Science, Tunku Abdul Rahman University, 53300 Setapak, Kuala Lumpur, Malaysia 2 School of Science and Technology, Universiti Malaysia Sabah, 88999 Kota Kinabalu, Sabah, Malaysia 3 Agrotechnology and Bioscience Division, Malaysian Nuclear Agency, Bangi, 43000 Kajang, Selangor, Malaysia Corresponding author : * [email protected] ABSTRACT Mirabilis jalapa L., commonly known as ‘four o’clock plant’ produces a strong, sweet smelling fragrance after the flowers open at late afternoon. It is a well known ornamental plant as the flowers of different colours can be found simultaneously on the same plant or an individual flower can be splashed with different colours. The colour-changing phenomenon is one of the unique characteristics of M. jalapa as it can display flowers with different colour when it matures. Apart from its ornamental value, it has also earned its place in herbal medicine practices around the world. Its array of biological activities continues to support its use worldwide for control of viruses, fungi and yeast. In this study, callus culture was initiated from the leaf explants of M. jalapa. The suitable strength of MS (Murashige and Skoog) nutrient media was determined and the effects of different types of auxins [4-amino-3,5,6-trichloro picolinic acid (picloram), 2,4- dichlorophenoxyacetic acid (2,4-D), -naphthaleneacetic acid (NAA)] and cytokinin [6-benzyl amino purine (BAP)] at 0.0 2.5, 5.0, 7.5, 10.0, and 20.0 M were investigated in order to determine the suitable callus induction and maintenance media. -

RED CLOVER This Plant May Become Weedy Or Invasive in Some Regions Or Habitats and May Displace Other Vegetation Trifolium Pratense L
Plant Guide Weediness RED CLOVER This plant may become weedy or invasive in some regions or habitats and may displace other vegetation Trifolium pratense L. if not properly managed. Consult your local NRCS Plant Symbol = TRPR2 Field Office, Cooperative Extension Service Office, or state natural resource or agriculture department Contributed by: Idaho Plant Materials regarding its status and use. Weed information is also available from the PLANTS web site at plants.usda.gov Description Trifolium pratense L., red clover, is an introduced biennial or short-lived perennial that grows as one of two types: medium (double-cut) or mammoth (single- cut). Red clover initiates growth from the plant crown. Plants have hollow, hairy stems and branches. Stem lengths of medium and mammoth types average 18 inches and 24-30 inches, respectively. Medium types have about 4 branches per stem; mammoth types have 6 branches per stem. USDA NRCS PLANTS Each leaf consists of a slender stalk bearing 3 leaflets. The taproot of red clover normally disintegrates in the second year and plants that Uses survive have developed secondary, extensively Red clover is the most widely grown of all the true branched roots. Flowers are borne in compact clovers and is the most important legume hay crop in clusters at the tips of the branches and are usually the northeastern United States. Red clover is rose-pink in color. The flower shape is similar to pea primarily used for hay, pasture, silage, and soil flowers except is more elongated and much smaller. improvement. It is a quick growing crop, easily Flower heads usually consist of up to 125 flowers. -

Conservation Plants Pocket ID Guide
About this guide.......... The purpose of this guide is to help you identify come commonly used conservation plants. Its color photos, line drawings and seed photos will help you make identifications. Also included are plant stand evaluation and recommended use charts. Keep this guide with you as long as you need it! Uses 1 2 Evaluating Stands Seeding success may not be obvious from visual observation. Use the chart below to determine whether your first-year stand is adequate. Lay a square-foot frame, or a circular frame with a 42.5-inch circumference on the ground. Count the number of seedlings within the frame, taking at least 10 counts for each 10 acres, in representative areas of the field. The table is based on pure stands; if a mixture of grass and legume is planted, reduce the numbers by the ratio of each species planted. Inadequate stands should be re-seeded. When a stand is judged to be between adequate and inadequate, it should be reevaluated after the second growing season. Warm-season grasses may need to be evaluated after the third growing season. 3 Seeds Early ID -- Seed is Key Grasses can be very difficult to identify in early growth stages. The seed may be the best identify aid. A seed retains its form and position in the ground through the seedling's early growth stages. To identify a seedling, carefully dig it up and compare it to photographs or actual seeds. 4 Plant Parts Conservation Plants The Elsberry Plant Materials Center The Elsberry Plant Materials Center is a 243-acre facility near Elsberry, Missouri. -

Comparative Analysis of Nodulation-Related Small Secreted Peptides Across Legume Species
Comparative analysis of nodulation-related small secreted peptides across legume species A DISSERTATION SUBMITTED TO THE FACULTY OF THE UNIVERSITY OF MINNESOTA BY Diana Trujillo IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY Nevin Young November 2017 © Diana Trujillo 2017 Acknowledgments I would like to thank all those who, one way or another, have made this work possible. First and foremost, thanks go to my advisor, Nevin Young, who helped shape my vision for this project and was always available to give me support or the necessary guidance in the right direction. I would also like to thank Kevin Silverstein who played a large role during the development of my LSE pipeline, and was a helpful mentor in bioinformatic matters. Thanks to the other members of my advisory committee, Peter Morrell, Michael Sadowsky, and Robert Stupar, who provided a fresh outlook and valuable advice that helped to improve this study. I would like to thank Joseph Guhlin and Peng Zhou who were always one step away when I had Unix concerns, and Shaun Curtin and Roxanne Denny who guided me through the technical aspects of growing or transforming Medicago. I am grateful that I had a strong network of friends and colleagues to discuss biology, coding, and life. To Allison Haaning, Beth Fallon, Christina Smith, Leland Werden, Derek Nedveck, and Eli Krumholz: thanks for sharing the journey. I would like to thank my mother, María Cecilia, who instilled my love of reading, truth-seeking, and plants. To my husband, Dylan Huss, thank you for always calling to see if I already ate (including as I typed these words). -

Biology of a New Virus Isolated from Lupinus Nootkatensis Plants in Alaska
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln U.S. Department of Agriculture: Agricultural Publications from USDA-ARS / UNL Faculty Research Service, Lincoln, Nebraska 2004 Biology of a new virus isolated from Lupinus nootkatensis plants in Alaska N. L. Robertson USDA, ARS Follow this and additional works at: https://digitalcommons.unl.edu/usdaarsfacpub Robertson, N. L., "Biology of a new virus isolated from Lupinus nootkatensis plants in Alaska" (2004). Publications from USDA-ARS / UNL Faculty. 1443. https://digitalcommons.unl.edu/usdaarsfacpub/1443 This Article is brought to you for free and open access by the U.S. Department of Agriculture: Agricultural Research Service, Lincoln, Nebraska at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Publications from USDA-ARS / UNL Faculty by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Plant Pathology (2004) 53, 569–576 Doi: 10.1046/j.0032-0862.2004.01058.x BlackwellBiology Publishing, Ltd. of a new virus isolated from Lupinus nootkatensis plants in Alaska N. L. Robertson USDA, ARS, Arctic Plant Germplasm Introduction and Research Project, 533 E. Fireweed Avenue, Palmer, AK 99645, USA A new virus named Nootka lupine vein-clearing virus (NLVCV) was isolated from Lupinus nootkatensis plants that were confined to a relatively small area in the Talkeetna mountains of south-central Alaska. Annual surveys (2000–03) con- sistently found leaf symptoms of pronounced vein clearing and mosaic on 3- to 4-week-old plants in late June. Spherical particles ≈30 nm in diameter were isolated from these leaves. Virions contained a single-stranded RNA of ≈4·0–4·2 kb and one species of capsid protein estimated to be ≈40 kDa.