Mediterránea 20
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

TERMES DE REFERENCE Services Techniques Pour Une Consultation
TERMES DE REFERENCE Services techniques pour une consultation pour l´élaboration, présentation et diffusion de baromètres de suivi des politiques publiques locales en matière de participation, égalité et jeunesse 0. INTRODUCTION Titre de l´intervention: Convenio «Appui aux processus de participation démocratique au Maroc» Nº 14-CO1-413. Financement: Agence Espagnole de Coopération Internationale pour le Développement (AECID). Budget global du programme : 2.595.000 €. Gestionnaires du programme: Espace Associatif (EA), Mouvement pour la Paix (MPDL) et Oxfam Intermón (OI). Durée du programme : 4 ans (2015-2018). Cette consultation sera réalisée dans le cadre du Programme (Convenio) « Appui aux processus de participation démocratique au Maroc », mis en œuvre par un consortium formé par l’Espace Associatif (EA), le Mouvement pour la Paix (MPDL) et Oxfam Intermón avec le financement de l'Agence Espagnole de Coopération Internationale au Développement (AECID). 1. CONTEXTE ET DETAILS DU PROGRAMME Les objectifs du programme sont les suivants : Objectif Général : Soutenir le processus de bonne gouvernance démocratique à travers le développement des capacités des acteurs locaux et la mise en place de mécanismes de participation citoyenne qui garantissent l’égalité entre les hommes et les femmes. Objectif Spécifique 1 : Favoriser la participation à travers la concertation et l’implication des différents acteurs dans la gestion des questions publiques locales. Objectif Spécifique 2 : Promouvoir l'égalité entre hommes et femmes dans les sphères publiques et privées au niveau local et national. Objectif Spécifique 3 : Améliorer les conditions de participation de la jeunesse dans la gestion des questions locales. Objectif Spécifique 4 : Améliorer et renforcer la capacité d’incidence des organisations de la société civile pour les convertir en une force de proposition et de promotion de la démocratie participative au niveau local, national et international. -

Leishmaniasis in Northern Morocco: Predominance of Leishmania Infantum Compared to Leishmania Tropica
Hindawi BioMed Research International Volume 2019, Article ID 5327287, 14 pages https://doi.org/10.1155/2019/5327287 Research Article Leishmaniasis in Northern Morocco: Predominance of Leishmania infantum Compared to Leishmania tropica Maryam Hakkour ,1,2,3 Mohamed Mahmoud El Alem ,1,2 Asmae Hmamouch,2,4 Abdelkebir Rhalem,3 Bouchra Delouane,2 Khalid Habbari,5 Hajiba Fellah ,1,2 Abderrahim Sadak ,1 and Faiza Sebti 2 1 Laboratory of Zoology and General Biology, Faculty of Sciences, Mohammed V University in Rabat, Rabat, Morocco 2National Reference Laboratory of Leishmaniasis, National Institute of Hygiene, Rabat, Morocco 3Agronomy and Veterinary Institute Hassan II, Rabat, Morocco 4Laboratory of Microbial Biotechnology, Sciences and Techniques Faculty, Sidi Mohammed Ben Abdellah University, Fez, Morocco 5Faculty of Sciences and Technics, University Sultan Moulay Slimane, Beni Mellal, Morocco Correspondence should be addressed to Maryam Hakkour; [email protected] Received 24 April 2019; Revised 17 June 2019; Accepted 1 July 2019; Published 8 August 2019 Academic Editor: Elena Pariani Copyright © 2019 Maryam Hakkour et al. Tis is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In Morocco, Leishmania infantum species is the main causative agents of visceral leishmaniasis (VL). However, cutaneous leishmaniasis (CL) due to L. infantum has been reported sporadically. Moreover, the recent geographical expansion of L. infantum in the Mediterranean subregion leads us to suggest whether the nonsporadic cases of CL due to this species are present. In this context, this review is written to establish a retrospective study of cutaneous and visceral leishmaniasis in northern Morocco between 1997 and 2018 and also to conduct a molecular study to identify the circulating species responsible for the recent cases of leishmaniases in this region. -

Pauvrete, Developpement Humain
ROYAUME DU MAROC HAUT COMMISSARIAT AU PLAN PAUVRETE, DEVELOPPEMENT HUMAIN ET DEVELOPPEMENT SOCIAL AU MAROC Données cartographiques et statistiques Septembre 2004 Remerciements La présente cartographie de la pauvreté, du développement humain et du développement social est le résultat d’un travail d’équipe. Elle a été élaborée par un groupe de spécialistes du Haut Commissariat au Plan (Observatoire des conditions de vie de la population), formé de Mme Ikira D . (Statisticienne) et MM. Douidich M. (Statisticien-économiste), Ezzrari J. (Economiste), Nekrache H. (Statisticien- démographe) et Soudi K. (Statisticien-démographe). Qu’ils en soient vivement remerciés. Mes remerciements vont aussi à MM. Benkasmi M. et Teto A. d’avoir participé aux travaux préparatoires de cette étude, et à Mr Peter Lanjouw, fondateur de la cartographie de la pauvreté, d’avoir été en contact permanent avec l’ensemble de ces spécialistes. SOMMAIRE Ahmed LAHLIMI ALAMI Haut Commissaire au Plan 2 SOMMAIRE Page Partie I : PRESENTATION GENERALE I. Approche de la pauvreté, de la vulnérabilité et de l’inégalité 1.1. Concepts et mesures 1.2. Indicateurs de la pauvreté et de la vulnérabilité au Maroc II. Objectifs et consistance des indices communaux de développement humain et de développement social 2.1. Objectifs 2.2. Consistance et mesure de l’indice communal de développement humain 2.3. Consistance et mesure de l’indice communal de développement social III. Cartographie de la pauvreté, du développement humain et du développement social IV. Niveaux et évolution de la pauvreté, du développement humain et du développement social 4.1. Niveaux et évolution de la pauvreté 4.2. -
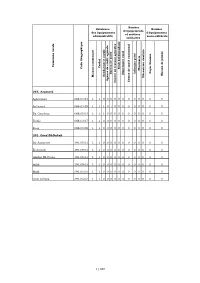
M a Is O N C O M M U N a Le C a Ïd a T G E N D a Rm E Rie Ro Y a Le A
Nombre Existence Nombre d'équipements des équipements d'équipements et services administratifs socio-culturels sanitaires Commune rurale Caïdat Code Géographique Souk Souk hebdomadaire Pharmacie Foyer féminin Infirmier privé Bureau deBureau poste Maison de jeunes Dispensaire rural Maison communale Mécanicien dentiste Gendarmerie royale Agence de crédit agricole Centre de santé communal Centre de travaux agricoles 066. Aousserd Aghouinite 066.03.03 1 1 0 0 0 0 0 0 0 0 0 0 0 0 Aousserd 066.03.05 1 1 1 0 1 0 0 0 0 0 0 0 0 0 Bir Gandouz 066.05.03 1 1 1 0 0 0 0 0 0 0 0 0 0 0 Tichla 066.03.07 1 1 0 0 0 0 0 0 0 0 0 0 0 0 Zoug 066.03.09 1 1 0 0 0 0 0 0 0 0 0 0 0 0 391. Oued Ed-Dahab Bir Anzarane 391.05.01 1 1 0 0 0 0 0 0 0 0 0 0 0 0 El Argoub 391.09.01 1 1 0 0 0 0 0 0 0 0 0 0 0 0 Gleibat EL Foula 391.05.03 1 1 0 0 0 0 0 0 0 0 0 0 0 0 Imlili 391.09.03 1 1 0 0 0 0 0 0 0 0 0 0 0 0 Mijik 391.05.05 1 1 0 0 0 0 0 0 0 0 0 0 0 0 Oum Dreyga 391.05.07 1 1 0 0 0 0 0 0 0 0 0 0 0 0 1/160 La commune Nombre d'établissements est Réseaux d'enseignement et de accessible d'infrastructure formation par Commune rurale Train Lycée Collège Autocar de développement ? Grand taxi Autre moyen Réseau d'électricité Réseau d'eau potable Ecole primaire satellite Ecole primaire centrale professionnelle publique Ecole coranique ou Msid Réseau d'assainissement La commune dispose-t-elle d'un plan Ecole primaire autonome Etablissement de formation 066. -

Activité 2: Aménagement Du Territoire Dans Un Contexte De Haut Risque
Document V1.0 Actions de la Fondation Ayesa au titre de l’activité 2 PRAVEMA Activité 2: Aménagement du territoire dans un contexte de haut risque Action 1: Définition du contexte de haut risque pour la population TABLE DES MATIERES 1. Introduction................................................................................. 1 2. Portée de l'étude. .......................................................................... 2 2.1. Critères de délimitation de la zone d'étude. .............................................. 3 2.2. Etude hydraulique simplifiée............................................................... 4 2.3. Zones du bassin historiquement inondables. ............................................. 6 2.4. Délimitation de la zone d'étude............................................................ 8 3. Organisation administrative................................................................ 9 3.1. Démographie ............................................................................. 10 3.2. Données socioéconomiques ............................................................. 15 4. Définition des contextes à haut risque d'inondation pour la population. ............. 23 5. Utilisation du sol.......................................................................... 29 5.1. Agglomérations principales. Larache et Ksar el Kébir. .................................. 36 5.2. Terrains inondables de l'Oued Loukkos entre le barrage Oued El Makhazine et son embouchure à Larache. ............................................................................... -

Le Délégué Provincial Objet : Modification
ROYAUME DU MAROC Ministère De L'Education Nationale, de L'Enseignement Supérieur, de la Formation Des Cadres et de la Recherche Scientifique SECRETARIAT D’ETAT CHARGE DE L’EDUCATION SCOLAIRE Académie Régionale d'Éducation et de la Formation Région Tanger/Tétouan Délégation Provinciale de Larache Service des Affaires Administratives et Financières Bureau du Budget Le délégué provincial Objet : Modification Réf : Avis d’Appel d'offres n° 01 /2010/Lar : du 17/02/2010 Suite aux offres n° : · 01/EXP/2010 relatif l’Achat de matières d’alimentation destinées aux internats des établissements du 2° Cycle fondamental et des établissements secondaires et techniques relevant de La délégation provinciale de Larache Une précision effectuée au niveau des articles des échantillons à déposer par les fournisseurs.(voir R.C -article 18 ) · 02/EXP/2010 relatif au Nettoyage des bâtiments administratifs et des sièges des établissements scolaires dépendants de la délégation provinciale de Larache Une modification effectuée au niveau de nombre des agents de nettoyage, comme suite : Il convient de dire : Au lieu de : ETABLISSEMENT A Nettoyer COMMUNE NOMBRE DE NOMBRE DE personnes personnes C. ABI LMAHASSINE KSAR EL KEBIR 05 10 C. ALLAL BEN ABDELLAH KSAR EL KEBIR 02 01 C. LAAOUAMRA LAAOUAMRA 05 10 C. RISSANA RISSANA 05 10 C.AZIZ BLAL BNI AROUS 02 01 C.BNI GUERFET BNI GUERFET 02 01 C.KOLLA KOLLA 02 01 C.MOHAMED BEN ABDELKRIM EL KHATTABI LARACHE 05 11 C.MOULAY ABDESSALAM BEN MECHIC TATOUFT 02 01 C.SAHEL SAHEL 02 01 C.TABARI KSAR EL KEBIR 02 01 C.ZOUADA ZOUADA 02 01 L. -

SP Larache.Pdf
ROYAUME DU MAROC Direction Régionale de Tanger Tétouan Al Hoceima RECENSEMENT GENERAL DE LA POPULATION ET DE L’HABITAT 2014 CARACTERISTIQUES DEMOGRAPHIQUES ET SOCIO-ECONOMIQUES PROVINCE DE LARACHE SERIE PROVINCIALE SOMMAIRE INTRODUCTION ............................................................................................................................................ 5 1 LES CARACTERISTIQUES DEMOGRAPHIQUES DE LA POPULATION ..................................... 9 1.1 Accroissement démographique et répartition spatiale de la population ........................................... 9 1.2 Population selon le sexe et l’âge .............................................................................................................. 9 1.3 Statut matrimonial de la population ........................................................................................................12 1.4 Fécondité .....................................................................................................................................................16 2 LES CARACTERISTIQUES DE L’EDUCATION .............................................................................. 21 2.1 L'aptitude à lire et à écrire .......................................................................................................................21 2.2 Scolarisation ................................................................................................................................................23 2.2.1 Scolarisation des enfants âgés de 7 à 12 ans ....................................................................................... -

La Conservation Des Sites Naturels Sacrés Au Maroc: Est-Elle Incompatible Avec Le Développement Socio-Économique? Índice
MEDITERRANEA SERIE DE ESTUDIOS BIOLÓGICOS 2009 Época II Nº 20 COMITÉ EDITORIAL: G.U. CARAVELLO S.G. CONARD A. FARINA A. FERCHICHI L. TAÏQUI Universitat d’Alacant Universidad de Alicante Departamento de Ecología. Facultad de Ciencias COMITÉ CIENTÍFICO: S. G. CONARD. USDA Forest Service. Riverside. U.S.A. A. FARINA. Lab. Ecologia del Paisaje. Museo Historia Natural. Aulla. Italia. A. FERCHICHI. I.R.A. Medenine. Túnez. G.U.CARAVELLO. Istituto di Igiene. Università di Padova. Italia. L. TAÏQUI. Université Abdelmalek Essaâdi. Tetuán. Marruecos. COMITÉ EDITORIAL: V. Peiró, J. Martín, G. López, E. Seva. DIRECCIÓN: Eduardo Seva. Dep. Ecología. Fac. de Ciencias. Universidad de Alicante. SECRETARÍA: Germán López. Dep. Ecología. Universidad de Alicante. EDITA: Servicio de Publicaciones. Universidad de Alicante. http://publicaciones.ua.es CORRESPONDENCIA: Departamento de Ecología. Fac. de Ciencias. Universidad de Alicante. Ap. 99 - 03080 Alicante. España. Teléfono de Secretaría: +34965903400, ext 2255 Fax: Rev. Mediterránea. Dep. Ecología. 96/5903464 I.S.S.N.: 0210-5004 Depósito Legal: A-1059-1984 Edición electrónica: Espagrafic Notas para los autores Los trabajos versarán sobre aspectos de ecología, recursos natu- rales, paisaje, gestión ambiental, en los ecosistemas de bioma me- diterráneo. Los manuscritos mecanografi ados a doble espacio y por una sola cara se enviarán a la dirección del Departamento de Ecología de la Universi- dad de Alicante, Ap. 99 (03080 Alicante, España) —Revista Mediter- ranea—. Los autores deberán enviar original y dos copias, así como en disquette compatible en programas de tratamiento de texto WORD. LENGUA: Redactados en español, inglés, francés o italiano. NOMBRE DE AUTORES: Apellidos y nombres sin abreviaciones. -

La Conservation Des Sites Naturels Sacrés Au Maroc: Est-Elle Incompatible Avec Le Développement Socio-Économique?
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/277202126 La conservation des sites naturels sacrés au Maroc: est-elle incompatible avec le développement socio-économique? Article in Mediterránea Serie de Estudios Biológicos · January 2009 DOI: 10.14198/MDTRRA2009.20.04 CITATIONS READS 12 1,337 3 authors, including: Lahcen Taiqui Eduardo Seva Abdelmalek Essaâdi University University of Alicante 36 PUBLICATIONS 319 CITATIONS 34 PUBLICATIONS 201 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Regional Knowledge Network on Water View project Social, Ecological and Agricultural Resilience in the face of climate CHange (SEARCH) View project All content following this page was uploaded by Lahcen Taiqui on 14 July 2015. The user has requested enhancement of the downloaded file. MEDITERRANEA SERIE DE ESTUDIOS BIOLÓGICOS 2009 Época II Nº 20 COMITÉ EDITORIAL: G.U. CARAVELLO S.G. CONARD A. FARINA A. FERCHICHI L. TAÏQUI Universitat d’Alacant Universidad de Alicante Departamento de Ecología. Facultad de Ciencias COMITÉ CIENTÍFICO: S. G. CONARD. USDA Forest Service. Riverside. U.S.A. A. FARINA. Lab. Ecologia del Paisaje. Museo Historia Natural. Aulla. Italia. A. FERCHICHI. I.R.A. Medenine. Túnez. G.U.CARAVELLO. Istituto di Igiene. Università di Padova. Italia. L. TAÏQUI. Université Abdelmalek Essaâdi. Tetuán. Marruecos. COMITÉ EDITORIAL: V. Peiró, J. Martín, G. López, E. Seva. DIRECCIÓN: Eduardo Seva. Dep. Ecología. Fac. de Ciencias. Universidad de Alicante. SECRETARÍA: Germán López. Dep. Ecología. Universidad de Alicante. EDITA: Servicio de Publicaciones. Universidad de Alicante. http://publicaciones.ua.es CORRESPONDENCIA: Departamento de Ecología. -

MOROCCO Cannabis Survey 2004
ROYAUME DU MAROC LE PREMIER MINISTRE AGENCE POUR LA PROMOTION ET LE DEVELOPPEMENT ECONOMIQUE ET SOCIAL DES PREFECTURES ET PROVINCES DU NORD DU ROYAUME Vienna International Centre, P.O. Box 500, A-1400 Vienna, Austria Tel: (+43 1) 26060-0, Fax: (+43 1) 26060-5866, www.unodc.org MOROCCO Cannabis Survey 2004 Executive Summary 2004 May 2005 Abbreviations APDN Agence pour la Promotion et le Développement Economique et Social des Préfectures et Provinces du Nord du Royaume CRTS Centre Royal de Télédétection spatiale DPAE Direction de la Programmation et des Affaires Economiques du Ministère de l’Agriculture, du Développement Rural et des Eaux et Forêts Dh Moroccan Dirham GPS Global Positioning System ICMP Illicit Crop Monitoring Programme LARATES Laboratoire de Recherches et d’Analyses Techniques et Scientifiques, Gendarmerie Royale UNODC United Nations Office on Drugs and Crime $ United States dollars Acknowledgments The following institutions and individuals contributed to the implementation of this survey and to the preparation of the present report: Government of Morocco: APDN : Direction des Affaires Administratives et Financières Direction des Opérations Département de la Coopération Internationale Département des Secteurs Productifs CRTS Centre Royal de Télédétection spatiale DPAE : Division des Statistiques et de l’Informatique LARATES : Département Analyses et Prélèvements, Gendarmerie Royale Association TARGA The implementation of this survey would not have been possible without the support of the local administration of the prefectures -

Code Géographique Du Maroc
Région: OUED ED-DAHAB-LAGOUIRA |_0_|_1_| Province ou Cercle Commune ou Arrondissement Préfecture Libellé Code Libellé Code Libellé Code Lagouira (M) 066.01.03 Aghouinite 066.03.03 Aousserd 066.03.05 Aousserd 066 Aousserd 066.03 Tichla 066.03.07 Zoug 066.03.09 Bir Gandouz 066.05 Bir Gandouz 066.05.03 Dakhla (M) 391.01.01 Bir Anzarane 391.05.01 Gleibat El Foula 391.05.03 Oued-Ed- Bir Anzarane 391.05 391 Mijik 391.05.05 Dahab Oum Dreyga 391.05.07 El Argoub 391.09.01 El Argoub 391.09 Imlili 391.09.03 Région:LAAYOUNE-BOUJDOUR-SAKIA EL HAMRA |_0_|_2_| Province ou Cercle Commune ou Arrondissement Préfecture Libellé Code Libellé Code Libellé Code Boujdour (M) 121.01.01 Gueltat Zemmour 121.03.01 Boujdour 121 Jraifia 121.03 Jraifia 121.03.03 Lamssid 121.03.05 El Marsa (M) 321.01.01 Laayoune (M) 321.01.03 Tarfaya (M) 321.01.05 Boukraa 321.03.01 Laayoune 321.03 Dcheira 321.03.03 Laayoune 321 Foum El Oued 321.03.05 Akhfennir 321.05.01 Daoura 321.05.03 Tarfaya 321.05 El Hagounia 321.05.05 Tah 321.05.07 Région: GUELMIM-ES-SEMARA |_0_|_3_| Province ou Cercle Commune ou Arrondissement Préfecture Libellé Code Libellé Code Libellé Code Assa (M) 071.01.01 Zag (M) 071.01.03 Aouint Lahna 071.03.01 Assa-Zag 071 Assa 071.03 Aouint Yghomane 071.03.03 Touizgui 071.03.05 Al Mahbass 071.05.01 Zag 071.05 Labouirat 071.05.03 Es-semara (M) 221.01.01 Amgala 221.03.01 Haouza 221.03.03 Es-Semara 221 Es-Semara 221.03 Jdiriya 221.03.05 Sidi Ahmed Laarouss 221.03.07 Tifariti 221.03.09 Bouizakarne (M) 261.01.01 Guelmim (M) 261.01.03 Aday 261.03.01 Ait Boufoulen 261.03.03 Amtdi -

Diagnostic Participatif Des Besoins Et Intérêts Des Femmes Et Des Jeunes Au Niveaux Des Régions De Tanger-Tétouan-Al Hoceïma Et De L’Oriental
Termes de Référence Diagnostic participatif des besoins et intérêts des femmes et des jeunes au niveaux des régions de Tanger-Tétouan-Al Hoceïma et de l’Oriental Projet Nº 18-CO1-1204: Appui à la consolidation des dynamiques de participation démocratique aux niveaux communal, provincial, régional et national pour contribuer à la réduction des inégalités socio-économiques au Maroc Financement : Agence Espagnole de Coopération Internationale pour le Développement (AECID). Durée du projet : 4 ans (2019-2022) SOMMAIRE 1. INTRODUCTION ET ANTÉCEDENTS 2. OBJECTIFS DE LA CONSULTATION 3. ACTEURS IMPLIQUÉS 4. MÉTHODOLOGIE ET PLAN DE TRAVAIL 5. ÉQUIPE DE CONSULTATION 6. DÉLAIS DE LA CONSULTATION 7. PRÉSENTATION DE L'OFFRE TECHNIQUE ET FINANCIERE Avec le financement de: 1. INTRODUCTION ET ANTÉCEDENTS Le document présent contient les termes de référence pour la réalisation d'un Diagnostic participatif des besoins et intérêts des femmes et des jeunes dans les régions de Tanger- Tétouan-Al-Hoceïma et la région de l’Oriental. Cette consultation fait partie de l'une des cinq études prévues dans le cadre du Projet Convenio1 Nº 18-CO1-1204: Appui à la consolidation des dynamiques de participation démocratique aux niveaux communal, provincial, régional et national pour contribuer à la réduction des inégalités socio-économiques au Maroc, co-exécuté par le consortium Mouvement pour la Paix (MPDL) et OXFAM au Maroc en collaboration avec leurs partenaires locaux. Cette intervention est la deuxième partie du Projet Convenio n ° 14-CO1-413 Appui aux processus de participation démocratique au Maroc, qui a été exécutée par les mêmes organisations entre 2015 et fin mars 2020.