Mountains As Evolutionary Arenas
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Quantifying Trends of Land Change in Qinghai-Tibet Plateau During 2001–2015
remote sensing Article Quantifying Trends of Land Change in Qinghai-Tibet Plateau during 2001–2015 Chao Wang, Qiong Gao and Mei Yu * Department of Environmental Sciences, University of Puerto Rico, Rio Piedras, San Juan, PR 00936, USA; [email protected] (C.W.); [email protected] (Q.G.) * Correspondence: [email protected]; Tel.: +1-787-764-0000 Received: 1 September 2019; Accepted: 17 October 2019; Published: 20 October 2019 Abstract: The Qinghai-Tibet Plateau (QTP) is among the most sensitive ecosystems to changes in global climate and human activities, and quantifying its consequent change in land-cover land-use (LCLU) is vital for assessing the responses and feedbacks of alpine ecosystems to global climate changes. In this study, we first classified annual LCLU maps from 2001–2015 in QTP from MODIS satellite images, then analyzed the patterns of regional hotspots with significant land changes across QTP, and finally, associated these trends in land change with climate forcing and human activities. The pattern of land changes suggested that forests and closed shrublands experienced substantial expansions in the southeastern mountainous region during 2001–2015 with the expansion of massive meadow loss. Agricultural land abandonment and the conversion by conservation policies existed in QTP, and the newly-reclaimed agricultural land partially offset the loss with the resulting net change of 5.1%. Although the urban area only expanded 586 km2, mainly at the expense of agricultural − land, its rate of change was the largest (41.2%). Surface water exhibited a large expansion of 5866 km2 (10.2%) in the endorheic basins, while mountain glaciers retreated 8894 km2 ( 3.4%) mainly in the − southern and southeastern QTP. -

Article-Lanzhou
13 by He Yuanqing, Yao Tandong, Cheng Guodong, and Yang Meixue Climatic records in a firn core from an Alpine temperate glacier on Mt. Yulong, southeastern part of the Tibetan Plateau Cold and Arid Regions Environmental and Engineering Research Institute, Chinese Academy of Sciences, Lanzhou 73000, China. Mt. Yulong is the southernmost glacier-covered area in Asian southwestern monsoon climate. Their total area is 11.61 km2. Eurasia, including China. There are 19 sub-tropical The glaciers resemble a group of flying dragons, giving Mt. Yulong temperate glaciers on the mountain, controlled by the (white dragon) as its name. Many explorers, tourists, poets and scientists have described southwestern monsoon climate. In the summer of 1999, Mt. Yulong from different points of view (Ward, 1924; Wissmann, a firn core, 10.10 m long, extending down to glacier ice, 1937). However, as they were unable to cross the extremely steep was recovered in the accumulation area of the largest mountain slopes and the forested area to the glacier above 4000 m a.s.l., some reported data were not correct. Most of the literature has glacier, Baishui No.1. Periodic variations of climatic focussed on the alpine landscape and the snow scenery, and there are signals above 7.8 m depth were apparent, and net accu- few accounts of the existing glaciers. Ren et al (1957) and Luo and mulation of four years was identified by the annual Yang (1963) first reported the distribution of these glaciers and of the late Pleistocene glacial landforms. To clarify the scale of glacia- oscillations of isotopic and ionic composition. -

Diversity and Geographical Pattern of Altitudinal Belts in the Hengduan Mountains in China
J. Mt. Sci. (2010) 7: 123–132 DOI: 10.1007/s11629-010-1011-9 Diversity and Geographical Pattern of Altitudinal Belts in the Hengduan Mountains in China YAO Yonghui, ZHANG Baiping*, HAN Fang, and PANG Yu Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China. E-mail: [email protected] * Corresponding author, e-mail: [email protected] © Science Press and Institute of Mountain Hazards and Environment, CAS and Springer-Verlag Berlin Heidelberg 2010 Abstract: This paper analyses the diversity and effect needs further study. In addition, the data spatial pattern of the altitudinal belts in the quality and data accuracy at present also affect to Hengduan Mountains in China. A total of 7 types of some extent the result of quantitative modeling and base belts and 26 types of altitudinal belts are should be improved with RS/GIS in the future. identified in the study region. The main altitudinal belt lines, such as forest line, the upper limit of dark Keywords: Hengduan Mountains; Altitudinal belt coniferous forest and snow line, have similar spectra; Exposure effect; Quadratic model latitudinal and longitudinal spatial patterns, namely, arched quadratic curve model with latitudes and concave quadratic curve model along longitudinal Introduction direction. These patterns can be together called as “Hyperbolic-paraboloid model”, revealing the complexity and speciality of the environment and Globally and generally, the upper and lower ecology in the study region. This result further limits of altitudinal belts vary (increase) from high validates the hypnosis of a common quadratic model latitudes to low latitudes, from continental for spatial pattern of mountain altitudinal belts peripheries to inland areas, and from very humid proposed by the authors. -

Essd-2020-71.Pdf
Discussions https://doi.org/10.5194/essd-2020-71 Earth System Preprint. Discussion started: 7 September 2020 Science c Author(s) 2020. CC BY 4.0 License. Open Access Open Data 1 Consolidating the Randolph Glacier Inventory and the Glacier 2 Inventory of China over the Qinghai-Tibetan Plateau and th 3 Investigating Glacier Changes Since the mid-20 Century 4 Xiaowan Liu1,2,3, Zongxue Xu1,2, Hong Yang3,4, Xiuping Li5, Dingzhi Peng1,2 5 1College of Water Sciences, Beijing Normal University, Beijing, 100875, China 6 2Beijiing Key Laboratory of Urban Hydrological Cycle and Sponge City Technology, Beijing, 100875, China 7 3Eawag, Swiss Federal Institute of Aquatic Science and Technology, 8600 Dübendorf, Switzerland 8 4Department of Environmental Science, MGU University of Basel, Petersplatz 1, 4001, Switzerland 9 5Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing, 100101, China 10 Correspondence to: Zongxue Xu ([email protected]) 11 Abstract. Glacier retreat in the Qinghai-Tibetan Plateau (QTP), the ‘third pole of the world’, has attracted the 12 attention of researchers worldwide. Glacier inventories in the 1970s and the 2000s provide valuable information 13 to infer changes in individual glaciers. However, individual glacier volumes are either missing, incomplete or have 14 large errors in these inventories, and thus, the use of these datasets to investigate changes in glaciers in QTP in the 15 past few decades has become a challenge, particularly in the context of climate change. In this study, individual 16 glacier volume data in the Randolph Glacier Inventory version 4.0 (RGI 4.0, 1970s) and the second Glacier 17 Inventory of China (GIC-Ⅱ, 2000s) are recalculated and consolidated using a slope-dependent algorithm based on 18 elevation datasets for the QTP. -

Downloaded from Brill.Com10/07/2021 12:04:04PM Via Free Access © Koninklijke Brill NV, Leiden, 2017
Amphibia-Reptilia 38 (2017): 517-532 A new moth-preying alpine pit viper species from Qinghai-Tibetan Plateau (Viperidae, Crotalinae) Jingsong Shi1,2,∗, Gang Wang3, Xi’er Chen4, Yihao Fang5,LiDing6, Song Huang7,MianHou8,9, Jun Liu1,2, Pipeng Li9 Abstract. The Sanjiangyuan region of Qinghai-Tibetan Plateau is recognized as a biodiversity hotspot of alpine mammals but a barren area in terms of amphibians and reptiles. Here, we describe a new pit viper species, Gloydius rubromaculatus sp. n. Shi, Li and Liu, 2017 that was discovered in this region, with a brief taxonomic revision of the genus Gloydius.The new species can be distinguished from the other congeneric species by the following characteristics: cardinal crossbands on the back, indistinct canthus rostralis, glossy dorsal scales, colubrid-like oval head shape, irregular small black spots on the head scales, black eyes and high altitude distribution (3300-4770 m above sea level). The mitochondrial phylogenetic reconstruction supported the validity of the new species and furthermore reaffirms that G. intermedius changdaoensis, G. halys cognatus, G. h. caraganus and G. h. stejnegeri should be elevated as full species. Gloydius rubromaculatus sp. n. was found to be insectivorous: preying on moths (Lepidoptera, Noctuidae, Sideridis sp.) in the wild. This unusual diet may be one of the key factors to the survival of this species in such a harsh alpine environment. Keywords: Gloydius rubromaculatus sp. n., insectivorous, new species, Sanjiangyuan region. Introduction leopards (Uncia uncia), wild yaks (Bos grun- niens) and Tibetan antelopes (Pantholops hodg- The Sanjiangyuan region (the Source of Three sonii) (Shen and Tan, 2012). -
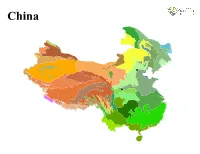
R Graphics Output
China China LEGEND Previously sampled Malaise trap site Ecoregion Alashan Plateau semi−desert North Tibetan Plateau−Kunlun Mountains alpine desert Altai alpine meadow and tundra Northeast China Plain deciduous forests Altai montane forest and forest steppe Northeast Himalayan subalpine conifer forests Altai steppe and semi−desert Northern Indochina subtropical forests Amur meadow steppe Northern Triangle subtropical forests Bohai Sea saline meadow Northwestern Himalayan alpine shrub and meadows Central China Loess Plateau mixed forests Nujiang Langcang Gorge alpine conifer and mixed forests Central Tibetan Plateau alpine steppe Ordos Plateau steppe Changbai Mountains mixed forests Pamir alpine desert and tundra Changjiang Plain evergreen forests Qaidam Basin semi−desert Da Hinggan−Dzhagdy Mountains conifer forests Qilian Mountains conifer forests Daba Mountains evergreen forests Qilian Mountains subalpine meadows Daurian forest steppe Qin Ling Mountains deciduous forests East Siberian taiga Qionglai−Minshan conifer forests Eastern Gobi desert steppe Rock and Ice Eastern Himalayan alpine shrub and meadows Sichuan Basin evergreen broadleaf forests Eastern Himalayan broadleaf forests South China−Vietnam subtropical evergreen forests Eastern Himalayan subalpine conifer forests Southeast Tibet shrublands and meadows Emin Valley steppe Southern Annamites montane rain forests Guizhou Plateau broadleaf and mixed forests Suiphun−Khanka meadows and forest meadows Hainan Island monsoon rain forests Taklimakan desert Helanshan montane conifer forests -

Long-Term Variations in Actual Evapotranspiration Over the Tibetan Plateau
Earth Syst. Sci. Data, 13, 3513–3524, 2021 https://doi.org/10.5194/essd-13-3513-2021 © Author(s) 2021. This work is distributed under the Creative Commons Attribution 4.0 License. Long-term variations in actual evapotranspiration over the Tibetan Plateau Cunbo Han1,2,3, Yaoming Ma1,2,4,5, Binbin Wang1,2, Lei Zhong6, Weiqiang Ma1,2,4, Xuelong Chen1,2,4, and Zhongbo Su7 1Land–Atmosphere Interaction and its Climatic Effects Group, State Key Laboratory of Tibetan Plateau Earth System and Resources Environment (TPESRE), Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing, China 2CAS Center for Excellence in Tibetan Plateau Earth Sciences, Chinese Academy of Sciences, Beijing, China 3Institute for Meteorology and Climate Research, Karlsruhe Institute of Technology, Karlsruhe, Germany 4College of Earth and Planetary Sciences, University of Chinese Academy of Sciences, Beijing, China 5Frontier Center for Eco-environment and Climate Change in Pan-third Pole Regions, Lanzhou University, Lanzhou, China 6Laboratory for Atmospheric Observation and Climate Environment Research, School of Earth and Space Sciences, University of Science and Technology of China, Hefei, China 7Faculty of Geo-Information Science and Earth Observation, University of Twente, Enschede, the Netherlands Correspondence: Y. Ma ([email protected]) Received: 30 October 2020 – Discussion started: 11 January 2021 Revised: 7 June 2021 – Accepted: 15 June 2021 – Published: 19 July 2021 Abstract. Actual terrestrial evapotranspiration (ETa) is a key parameter controlling land–atmosphere interac- tion processes and water cycle. However, spatial distribution and temporal changes in ETa over the Tibetan Plateau (TP) remain very uncertain. Here we estimate the multiyear (2001–2018) monthly ETa and its spatial distribution on the TP by a combination of meteorological data and satellite products. -

An Endemic Rat Species Complex Is Evidence of Moderate
www.nature.com/scientificreports OPEN An endemic rat species complex is evidence of moderate environmental changes in the Received: 14 June 2016 Accepted: 13 March 2017 terrestrial biodiversity centre of Published: 10 April 2017 China through the late Quaternary Deyan Ge1,*, Liang Lu2,*, Jilong Cheng1,3, Lin Xia1, Yongbin Chang1, Zhixin Wen1, Xue Lv1,3, Yuanbao Du1,3, Qiyong Liu2 & Qisen Yang1 The underlying mechanisms that allow the Hengduan Mountains (HDM), the terrestrial biodiversity centre of China, to harbour high levels of species diversity remain poorly understood. Here, we sought to explore the biogeographic history of the endemic rat, Niviventer andersoni species complex (NASC), and to understand the long-term persistence of high species diversity in this region. In contrast to previous studies that have proposed regional refuges in eastern or southern of the HDM and emphasized the influence of climatic oscillations on local vertebrates, we found that HDM as a whole acted as refuge for the NASC and that the historical range shifts of NASC mainly occurred in the marginal regions. Demographic analyses revealed slight recent population decline in Yunnan and south- eastern Tibet, whereas of the populations in Sichuan and of the entire NASC were stable. This pattern differs greatly from classic paradigms of temperate or alpine and holarctic species. Interestingly, the mean elevation, area and climate of potential habitats of clade a (N. excelsior), an alpine inhabitant, showed larger variations than did those of clade b (N. andersoni), a middle-high altitude inhabitant. These species represent the evolutionary history of montane small mammals in regions that were less affected by the Quaternary climatic changes. -

Phylogeography of Prunus Armeniaca L. Revealed by Chloroplast DNA And
www.nature.com/scientificreports OPEN Phylogeography of Prunus armeniaca L. revealed by chloroplast DNA and nuclear ribosomal sequences Wen‑Wen Li1, Li‑Qiang Liu1, Qiu‑Ping Zhang2, Wei‑Quan Zhou1, Guo‑Quan Fan3 & Kang Liao1* To clarify the phytogeography of Prunus armeniaca L., two chloroplast DNA fragments (trnL‑trnF and ycf1) and the nuclear ribosomal DNA internal transcribed spacer (ITS) were employed to assess genetic variation across 12 P. armeniaca populations. The results of cpDNA and ITS sequence data analysis showed a high the level of genetic diversity (cpDNA: HT = 0.499; ITS: HT = 0.876) and a low level of genetic diferentiation (cpDNA: FST = 0.1628; ITS: FST = 0.0297) in P. armeniaca. Analysis of molecular variance (AMOVA) revealed that most of the genetic variation in P. armeniaca occurred among individuals within populations. The value of interpopulation diferentiation (NST) was signifcantly higher than the number of substitution types (GST), indicating genealogical structure in P. armeniaca. P. armeniaca shared genotypes with related species and may be associated with them through continuous and extensive gene fow. The haplotypes/genotypes of cultivated apricot populations in Xinjiang, North China, and foreign apricot populations were mixed with large numbers of haplotypes/ genotypes of wild apricot populations from the Ili River Valley. The wild apricot populations in the Ili River Valley contained the ancestral haplotypes/genotypes with the highest genetic diversity and were located in an area considered a potential glacial refugium for P. armeniaca. Since population expansion occurred 16.53 kyr ago, the area has provided a suitable climate for the population and protected the genetic diversity of P. -

China: a Rich Flora Needed of Urgent Conservationprovided by Digital.CSIC
Orsis 19, 2004 49-89 View metadata, citation and similar papers at core.ac.uk brought to you by CORE China: a rich flora needed of urgent conservationprovided by Digital.CSIC López-Pujol, Jordi GReB, Laboratori de Botànica, Facultat de Farmàcia, Universitat de Barcelona, Avda. Joan XXIII s/n, E-08028, Barcelona, Catalonia, Spain. Author for correspondence (E-mail: [email protected]) Zhao, A-Man Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, The People’s Republic of China. Manuscript received in april 2004 Abstract China is one of the richest countries in plant biodiversity in the world. Besides to a rich flora, which contains about 33 000 vascular plants (being 30 000 of these angiosperms, 250 gymnosperms, and 2 600 pteridophytes), there is a extraordinary ecosystem diversity. In addition, China also contains a large pool of both wild and cultivated germplasm; one of the eight original centers of crop plants in the world was located there. China is also con- sidered one of the main centers of origin and diversification for seed plants on Earth, and it is specially profuse in phylogenetically primitive taxa and/or paleoendemics due to the glaciation refuge role played by this area in the Quaternary. The collision of Indian sub- continent enriched significantly the Chinese flora and produced the formation of many neoen- demisms. However, the distribution of the flora is uneven, and some local floristic hotspots can be found across China, such as Yunnan, Sichuan and Taiwan. Unfortunately, threats to this biodiversity are huge and have increased substantially in the last 50 years. -

Interdecadal Variation of Precipitation Over the Hengduan Mountains During Rainy Seasons
15 JUNE 2019 D O N G E T A L . 3743 Interdecadal Variation of Precipitation over the Hengduan Mountains during Rainy Seasons a,b,c a,b b,d b,d,e DANHONG DONG, WEICHEN TAO, WILLIAM K. M. LAU, ZHANQING LI, a,c,f,g a,h GANG HUANG, AND PENGFEI WANG a State Key Laboratory of Numerical Modeling for Atmospheric Sciences and Geophysical Fluid Dynamics, Institute of Atmospheric Physics, Chinese Academy of Sciences, Beijing, China b Earth System Science Interdisciplinary Center, University of Maryland, College Park, College Park, Maryland c University of Chinese Academy of Sciences, Beijing, China d Department of Atmospheric and Oceanic Science, University of Maryland, College Park, College Park, Maryland e State Key Laboratory of Remote Sensing Science, College of Global Change and Earth System Science, Beijing Normal University, Beijing, China f Laboratory for Regional Oceanography and Numerical Modeling, Qingdao National Laboratory for Marine Science and Technology, Qingdao, China g Joint Center for Global Change Studies, Beijing, China h Center for Monsoon System Research, Institute of Atmospheric Physics, Chinese Academy of Sciences, Beijing, China (Manuscript received 8 October 2018, in final form 20 February 2019) ABSTRACT The present study investigates the interdecadal variation of precipitation over the Hengduan Mountains (HM) during rainy seasons from various reanalysis and observational datasets. Based on a moving t test and Lepage test, an obvious rainfall decrease is identified around 2004/05. The spatial distribution of the rainfall changes exhibits large and significant precipitation deficits over the southern HM, with notable anomalous lower-level easterly divergent winds along the southern foothills of the Himalayas (SFH). -

Songtsam's Shangrila to Lhasa
SONGTSAM’S SHANGRILA TO LHASA EXPLORATORY ROAD TRIP SCENIC HIGHWAY G318; THE ROAD LESS TRAVELED IN THE HIDDEN KINGDOM OF EASTERN TIBET by WILLIAM D BOEHM PART 1 SHANGRI-LA YUNNAN Songtdsam landrovers and photographers at Meil, Yunnan beginning the first foreign Trip into Kham Easter Tibet Autonomous Region from Shagril-La Yunnan to Lhasa via Highway G318, a route that includes part of the traditional Tea Horse Trail Beginning in the Song Dynasty In the seventh century, the elite of Tibet’s feudal Buddhist theocracy discovered Pu er tea found in Sichuan and SW Yunnan. Supplying its growing demand meant forging a network of trails across China for both porters and mule caravans through steep mountainous forests, over 5,000 m (16,400 ft) passes, along deep gorges, and across unnavigable rivers traversed by rope and cable. The journey was long, 4000 km (2400 mi) with the valuable tea carried by porters, or mule caravans. In return, China needed the Tibetan sturdy war horse to fight the northern invaders. The trade route became known as the Tea Horse Road, the Cha Ma Dao. It still exists with some of it forgotten along ancient stone trails found in forests, some of it paved over beneath modern highways. A few of the men who still live have shared about survival of often bitter lives on the trails. They were the leaders of the mule caravans, the Ma Guo Tou, a culture of men who braved snow storms, bandits, and treacherous river crossings through one of the longest trade routes of the ancient world.