Understanding Sprint-Cycling Performance: the Integration of Muscle Power, Resistance, and Modeling
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Issue 47 B H P C Newsletter - Issue 47 Front Cover: Andy Pegg in Action at Eastway Photo: Brian Dalton
The British Human B H P C Power Club ...much better than Thomas the Tank Engine! Further Beatles Collaborations Ruled Out As Ringo Finds New Career Opportunity Winter 1996 Issue 47 B H P C Newsletter - Issue 47 Front Cover: Andy Pegg in action at Eastway Photo: Brian Dalton Contents Editorial Dave Larrington 3 Events '97 mostly gNick 7 Letter Roy MacDonald 9 Letter Robin Downes 10 Letter Hugh Haines 10 Rolling Resistance Colin Porter, John Kingsbury, 12 Aarn Tate, Roy MacDonald The Scouse School Of Aerodynamics Nigel Sleigh 16 Secretarial Stuff Steve Donaldson 17 Leicester - Lancaster Sherri Prisk 19 Results '96 - Bumper Edition Various Artists 24 Suppliers & Wants 40 Recumbent Christmas Tina Larrington 44 Objectives: The British Human Power Club was formed to foster all aspects of human-powered vehicles - air, land & water - for competitive, recreational and utility activities, to stimulate innovation in design and development in all spheres of HPV's, and to promote and to advertise the use of HPV's in a wide range of activities. And to have a good laugh while doing it. OFFICERS Chairman & Press Officer Mike Burrows 16 Thunder Lane ( Work 01603 721000 Thorpe, Norfolk ( Home 01603 32142 Competition Secretary gNick Green 267 Tixall Road ( Home 01785 223576 Stafford, ST16 3XS Secretary Steve Donaldson 21 Glenholme Avenue ( Home 01224 722514 Dyce, Aberdeen AB21 7FF Treasurer & Membership Dennis Adcock all cheques to be 29 Chequers Hill ( Home 01494 721088 made out to the Amersham, Bucks BHPC HP7 9DQ Newsletter Editor Dave Larrington 166 Higham Hill Road ( Home 0181 531 4496 London E17 6EJ E-mail: [email protected] Note new Secretary & Newsletter Editor.. -

Intro to Track Cycling
Introduction to Track Cycling What to Expect and How to Upgrade Photo: Snowy Mountain Photography Track cycling history ¨ Track racing dates back to the late 1800s and 6-day racing events Velodrome basics ¨ Velodromes can range from less than 200 meters to over 500 meters in length ¨ Wood, concrete, and asphalt 2012 London Olympic Velodrome are common surface materials ¨ Current Olympic velodrome standard is a wood indoor 250 meter velodrome with banking of around 45 degrees ¨ Ed Rudolph Velodrome (aka Northbrook) is a 382 meter asphalt velodrome with banking of around 20 degrees Ed Rudolph Velodrome What are the colored lines on the track? ¨ The ”blue band” or “cote d’azur” marks the track’s inside boundary. Racers may not ride on or below this band. The area below the blue band extending to the grass is called the “apron.” ¨ The black “measurement line” is used to measure the distance around the track. When doing pursuits or time trials, use this line as a guide. ¨ The red “sprinter’s line” defines the border of the sprint lane. The leading rider in this lane is said to “own the lane” and may only be passed by a rider going over on the right. NO PASSING BELOW RIDERS IN THE SPRINTERS LANE. Additionally once a sprint is engaged, a racer who is leading and in the sprinter’s lane can not leave it. ¨ The uppermost blue line is the “stayer’s line” or the relief line. It marks the boundary between faster and slower traffic, with the faster riders below the line and the slower “relief” riders above the line. -

2017 USA Cycling Rulebook
Chapter 7 National Championships 149 7. Championships The following sections apply to National Championships in the disciplines and age groups specified. See section 7J for specific differences between National Championships and State Championships 7A. Organization 7A1. The rights to organize National Championships may be awarded to local Race Directors who meet the requirements established by the CEO. 7A2. Massed start races with fewer than 10 participants may be combined with another category at the discretion of USA Cycling and the Chief Referee with riders being scored separately at the end of the event. 7A3. In National Championship events, the defending National Champion (in that event) shall be given highest priority in call-ups except if the event is run under UCI rules. In track events where heats are required, the defending National Champion must compete in the heats. 7A4. Para-cycling National Championships for cyclists with disabilities may be held in conjunction with other national championships. Classifications of para-cycling riders and regulations of competition will follow the Functional Classification System outlined by the UCI. 7B. National Championship Eligibility 7B1. National Championships are open only to riders who hold USA Cycling rider annual licenses or recognized license from a UCI affiliated federation, and meet other qualifications stated in these rules. (a) National Championships for Junior 17-18, Under 23, and Elites may only be entered by US Citizens with a USA racing nationality. (b) Regardless of any general rule pertaining to National Championship eligibility, any National Championship that is a direct qualifier for the World Championships or Olympic Games may only be entered by riders who are eligible under 150 international regulations to enter those events as part of the U.S. -

2017 USA Cycling Rulebook
2017RULE BOOK Welcome! On behalf of USA Cycling, we hope that you are looking forward to a new year of bike racing. We are glad that you are a member and hope that you will find many opportunities to enjoy bike racing of all kinds. Good luck with your racing! Rev. 1/20/17 Modified 1E6(d) chart to correctly reflect regulation Added header to 1J6 Modified 1H10(c) to reflect Cat. 5 women Minor formatting changes Rev. 2/21/17 Modified 1A3(c) to fix issue with one-day MTB licenses. Cover Photos: Men’s Road: Casey B. Gibson Women’s MTB: Phil Beckman/PB Creative Men’s CX: Weldon Weaver Women’s Track: Rob Jones/CanadianCyclist.com 1 This Rulebook is published by USA Cycling. It is organized as follows: Chapter 1 – General Regulations Chapter 2 – Track Chapter 3 – Road and Stage Racing Chapter 4 – Cyclo-cross Chapter 5 – Mountain Bike Chapter 6 – Collegiate Chapter 7 – Championships Chapter 8 -- Discipline Chapter 9 – Records Chapter 10 -- Gran Fondo Appendices Glossary Copies may be downloaded from the USAC website at www.usacycling.org. Officials are sent a hard copy. Other members may request a hard copy by sending a self- addressed mailing label and note that says "rulebook" to the address below: USA Cycling/ Attn: Technical Director 210 USA Cycling Point, Suite 100 Colorado Springs, CO 80919 Schedule of fees, USA Cycling Bylaws, Policies, Records, and Results of National Championships may be found online at www.usacycling.org Unfortunately, the English language does not have a neutral gender personal pronoun. -

International OMNIUM
International OMNIUM MEN 1) Flying Lap (against the clock) The International Omnium event 2) 30 km Points Race (15 km for junior men) is a multi-race event for individuals in track 3) Elimination cycling. Historically the omnium has had a 4) 4 km Individual Pursuit (3 km for junior men) variety of formats. Currently, and for the 2012 5) Scratch Race London Olympic Games, the omnium as defined 6) 1km Time trial by the Union Cycliste Internationale (UCI) and consists of six events (both timed individual *Timed events are conducted individually while events and massed start pack races) for men the rest are pack style races. and for women that are conducted over two consecutive days. Ideally, the Omnium event showcases the best all-round, consistent rider -- speed, endurance and savvy race intelligence make up an International Omnium champion. Points are awarded in reverse order for each event within the omnium. The rider who finishes first in an event receives one point, the second rider will gets two points and so on down the placings. The winner is the rider with the lowest total points. If two riders are tied on points, the combined time of the three time trials will be the tie breaker to determine final placing. Also, riders must complete every event in the omnium. So if WOMEN a rider were to crash in an early segment and not 1) Flying Lap (against the clock) make it to the finish, they would be eliminated 2) 20 km Points Race (10 km for junior women) from continuing on in the next portion. -

Modeling Sprint Cycling Using Field-Derived Parameters and Forward Integration
Modeling Sprint Cycling Using Field-Derived Parameters and Forward Integration JAMES C. MARTIN1, A. SCOTT GARDNER2,3, MARTIN BARRAS2, and DAVID T. MARTIN2 1The University of Utah, Department of Exercise and Sport Science, Salt Lake City, UT; 2Australian Institute of Sport, Canberra, Australian Capital Territory, AUSTRALIA; and 3Queensland Academy of Sport, Brisbane, Queensland, AUSTRALIA ABSTRACT MARTIN, J. C., A. S. GARDNER, M. BARRAS, and D. T. MARTIN. Modeling Sprint Cycling Using Field-Derived Parameters and Forward Integration. Med. Sci. Sports Exerc.,Vol. 38, No. 3, pp. 592–597, 2006. We previously reported that a mathematical model could accurately predict steady-state road-cycling power when all the model parameters were known. Application of that model to competitive cycling has been limited by the need to obtain accurate parameter values, the non–steady-state nature of many cycling events, and because the validity of the model at maximal power has not been established. Purpose: We determined whether modeling parameters could be accurately determined during field trials and whether the model could accurately predict cycling speed during maximal acceleration using forward integration. Methods: First, we quantified aerodynamic drag area of six cyclists using both wind tunnel and field trials allowing for these two techniques to be compared. Next, we determined the aerodynamic drag area of three world-class sprint cyclists using the field-test protocol. Track cyclists also performed maximal standing-start time trials, during which we recorded power and speed. Finally, we used forward integration to predict cycling speed from power–time data recorded during the maximal trials allowing us to compare predicted speed with measured speed. -

Example: Track Session Plan 3
ATTACHMENT 10 EXAMPLE: TRACK SESSION PLAN 3 COACH DATE TIME VENUE <<NAME>> <<DAY/MONTH/2019>> <<ONE HOUR>>pm TE AWAMUTU VELODROME EQUIPMENT REQUIRED RIDERS Cones [Skill level – Intermediate experienced] Motorbike <<RIDER’S NAME>> (AGE) – ( ) <<PHONE NUMBER>> Riders to bring their own gearing/tool kits <<RIDER’S NAME>> (AGE) – ( ) <<PHONE NUMBER>> <<RIDER’S NAME>> (AGE) – ( ) <<PHONE NUMBER>> SESSION FORMAT SESSION GOALS 15 mins Warm up Setting parameters for this and future track sessions 25 mins Set activities (Flying and Standing Laying the base foundations for flying and standing starts) starts 20 mins Skills session – using full banking Introduction to motor pacing when taking turns / changing 20 mins Motor pacing 10 mins Warm Down TECHNIQUES, ACTIVITIES, GAMES AND PHYSICAL TRAINING POTENTIAL HAZARDS, SOLUTIONS OR ACCIDENTS WARM UP: Please refer to the risk Meet and greet 15 minutes before designated start time of session – assessment plan for in depth get settled have a review of last session, discuss coaching points from analysis – but the main physical that session then have a chat about today’s session risks are: Both riders warm up together rolling round above blue line then faster 3 x drains @ 25kph then @ 30kph then @ 35kph (5 mins at each speed) 1 x broken concrete (identified), SET DRILLS gates to track needing to be 2 x flying sprints @ 85% (from 2000m mark to finish line) – to follow on closed and directly from warm up…reinforce skill session from last wee using a motorbike for • x standing starts @100m (from the 1000m – finish line) motorpacing • x standing 200m starts NOTES (All times are recorded and summary sheet sent to riders to enter into While packing up – have a chat their training logs after session) about the session SKILL To reinforce the skills required Using the banking to take turns / effect change of rider at the front of for standing starts, will involve bunch more speed work behind the • Discuss with riders on grass first and walk through what we are motorbike and will focus on trying to achieve. -
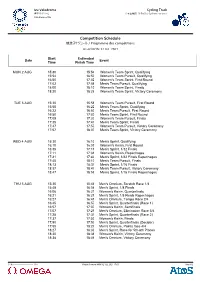
Competition Schedule
Izu Velodrome Cycling Track 伊豆ベロドローム 自転車競技(トラック) / Cyclisme sur piste Vélodrome d'Izu Competition Schedule 競技スケジュール / Programme des compétitions As of MON 12 JUL 2021 at HH:MM Start Estimated Date Event Time Finish Time MON 2 AUG 15:30 15:54 Women's Team Sprint, Qualifying 15:54 16:50 Women's Team Pursuit, Qualifying 16:50 17:02 Women's Team Sprint, First Round 17:02 17:58 Men's Team Pursuit, Qualifying 18:00 18:12 Women's Team Sprint, Finals 18:20 18:28 Women's Team Sprint, Victory Ceremony TUE 3 AUG 15:30 15:58 Women's Team Pursuit, First Round 15:58 16:22 Men's Team Sprint, Qualifying 16:22 16:50 Men's Team Pursuit, First Round 16:50 17:02 Men's Team Sprint, First Round 17:05 17:33 Women's Team Pursuit, Finals 17:35 17:47 Men's Team Sprint, Finals 17:47 17:57 Women's Team Pursuit, Victory Ceremony 17:57 18:07 Men's Team Sprint, Victory Ceremony WED 4 AUG 15:30 16:10 Men's Sprint, Qualifying 16:10 16:35 Women's Keirin, First Round 16:35 17:11 Men's Sprint, 1/32 Finals 17:11 17:31 Women's Keirin, Repechages 17:31 17:43 Men's Sprint, 1/32 Finals Repechages 17:45 18:13 Men's Team Pursuit, Finals 18:13 18:37 Men's Sprint, 1/16 Finals 18:37 18:47 Men's Team Pursuit, Victory Ceremony 18:47 18:59 Men's Sprint, 1/16 Finals Repechages THU 5 AUG 15:30 15:48 Men's Omnium, Scratch Race 1/4 15:48 16:06 Men's Sprint, 1/8 Finals 16:06 16:21 Women's Keirin, Quarterfinals 16:21 16:27 Men's Sprint, 1/8 Finals Repechages 16:27 16:45 Men's Omnium, Tempo Race 2/4 16:45 16:57 Men's Sprint, Quarterfinals (Race 1) 16:57 17:07 Women's Keirin, Semifinals 17:07 -

Using Field Based Data to Model Sprint Track Cycling Performance Hamish A
Ferguson et al. Sports Medicine - Open (2021) 7:20 https://doi.org/10.1186/s40798-021-00310-0 REVIEW ARTICLE Open Access Using Field Based Data to Model Sprint Track Cycling Performance Hamish A. Ferguson1* , Chris Harnish2 and J. Geoffrey Chase1 Abstract Cycling performance models are used to study rider and sport characteristics to better understand performance determinants and optimise competition outcomes. Performance requirements cover the demands of competition a cyclist may encounter, whilst rider attributes are physical, technical and psychological characteristics contributing to performance. Several current models of endurance-cycling enhance understanding of performance in road cycling and track endurance, relying on a supply and demand perspective. However, they have yet to be developed for sprint-cycling, with current athlete preparation, instead relying on measures of peak-power, speed and strength to assess performance and guide training. Peak-power models do not adequately explain the demands of actual competition in events over 15-60 s, let alone, in World-Championship sprint cycling events comprising several rounds to medal finals. Whilst there are no descriptive studies of track-sprint cycling events, we present data from physiological interventions using track cycling and repeated sprint exercise research in multiple sports, to elucidate the demands of performance requiring several maximal sprints over a competition. This review will show physiological and power meter data, illustrating the role of all energy pathways in sprint performance. This understanding highlights the need to focus on the capacity required for a given race and over an event, and therefore the recovery needed for each subsequent race, within and between races, and how optimal pacing can be used to enhance performance. -

Characteristics of Track Cycling
Sports Med 2001; 31 (7): 457-468 REVIEW ARTICLE 0112-1642/01/0007-0457/$22.00/0 © Adis International Limited. All rights reserved. Characteristics of Track Cycling Neil P. Craig1 and Kevin I. Norton2 1 Australian Institute of Sport, Track Cycling Unit Adelaide, South Australia, Australia 2 School of Physical Education, Exercise and Sport Studies, University of South Australia, Adelaide, South Australia, Australia Contents Abstract . 457 1. Track Cycling Events . 458 1.1 Energetics of Track Events . 458 2. Physical and Physiological Characteristics of Track Cyclists . 459 2.1 Body Shape, Size and Composition . 459 2.2 Maximal Oxygen Consumption . 461 2.3 Blood Lactate Transition Thresholds . 461 2.4 Anaerobic Capacity . 461 3. Competition Power Output . 462 3.1 200m Sprint . 462 3.2 1000m Time Trial . 463 3.3 4000m Team Pursuit . 463 3.4 4000m Individual Pursuit . 464 3.5 Madison . 464 4. Programme Design and Monitoring . 465 5. Conclusion . 466 Abstract Track cycling events range from a 200m flying sprint (lasting 10 to 11 seconds) to the 50km points race (lasting ≈1 hour). Unlike road cycling competitions where most racing is undertaken at submaximal power outputs, the shorter track events require the cyclist to tax maximally both the aerobic and anaerobic (oxygen independent) metabolic pathways. Elite track cyclists possess key physical and physiological attributes which are matched to the specific requirements of their events: these cyclists must have the appropriate genetic predisposition which is then maximised through effective training interventions. With advances in tech- nology it is now possible to accurately measure both power supply and demand variables under competitive conditions. -

Olympic Sprint, Or Team Sprint
Spectators Summary of Track Cycling Events: (See the USCF rule book or http://www.usacycling.org/ for all the details.) Mass Start or "Scratch" Race: Not surprisingly, all the riders in a Mass Start race start at the same time. The riders all cover the same distance, with the winner being the first rider to cross the finish line at the end of that distance. Although speed is important, tactics and teamwork are equally vital. Groups of riders often take an early lead, and then work together to increase it while their teammates try to block and slow down the "field." On a small track, the leaders may gain an entire lap on the other riders and then join in with the main group again. Points Race: A variation of the Mass Start race, points are awarded to the top placing riders in a series of sprints contested at various intervals during the race. The winner of a Points Race is not necessarily the first to cross the finish line, but rather the rider who has accumulated the most points during the race. Win-And-Out: A variation on the Scratch race where 1st place is decided on the final lap, however, only the winner is finished. In order to secure 2nd place that rider must be first across the line on the next lap. 3rd place is decided on the lap after that. Typically all other places are also decided by this third and final sprint. This makes for interesting tactics. It can be a very hard race if a rider tries gives their all to win only to be forced to continue and try again the next lap. -

Cycling Ibgn
PERFORMANCE CYCLING CONDITIONING A NEWSLETTER DEDICATED TO IMPROVING CYCLISTS www.performancecondition.com/cycling Periodization of Sprint Cycling with Considerations for Endurance Cyclists Clay Worthington, USA Cycling Assistant Coach Sprint Track, Colorado Springs, CO Clay spent two years as a scholarship coaching intern, an additional 6 months as assistant coach of sprint track for USAC, and is in his first season as Head Coach of Sprint Track. His responsibilities include implementation of training plans for sprint track at training sessions and perform necessary testing as required. He has also coached at multiple camps/trips (e.g., track en- durance, track sprint, road endurance, Southern and Palo Saeco Games-Trinidad). He is a USAC Level 1 Coach and a Cat 3 road and track licensed cyclist. He has a Masters of Science degree in kinesiology at Midwestern State University. BGN INT n cycling, the age-old question is quality versus quantity (i.e. intensity v. volume). If you look at the power numbers XTP there’s a wide degree for variance in power output. With women, the greatest power output for world-class athletes is be- MSR I tween 1300 and 1500 watts. With men, they are able to produce in the neighborhood of 2300 to 2500 watts (Figure 1). MTB Sprint athletes will try to produce these numbers on a regular basis—they try to practice at this maximum intensity. They do this from starts or as part of accelerations. With endurance athletes, if you put them on a bike at the end of a race the best any of them could produce would be anywhere from 1600 to 1700 watts.