Flood Pulses and River Ecosystem Robustness
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(P 117-140) Flood Pulse.Qxp
117 THE FLOOD PULSE CONCEPT: NEW ASPECTS, APPROACHES AND APPLICATIONS - AN UPDATE Junk W.J. Wantzen K.M. Max-Planck-Institute for Limnology, Working Group Tropical Ecology, P.O. Box 165, 24302 Plön, Germany E-mail: [email protected] ABSTRACT The flood pulse concept (FPC), published in 1989, was based on the scientific experience of the authors and published data worldwide. Since then, knowledge on floodplains has increased considerably, creating a large database for testing the predictions of the concept. The FPC has proved to be an integrative approach for studying highly diverse and complex ecological processes in river-floodplain systems; however, the concept has been modified, extended and restricted by several authors. Major advances have been achieved through detailed studies on the effects of hydrology and hydrochemistry, climate, paleoclimate, biogeography, biodi- versity and landscape ecology and also through wetland restoration and sustainable management of flood- plains in different latitudes and continents. Discussions on floodplain ecology and management are greatly influenced by data obtained on flow pulses and connectivity, the Riverine Productivity Model and the Multiple Use Concept. This paper summarizes the predictions of the FPC, evaluates their value in the light of recent data and new concepts and discusses further developments in floodplain theory. 118 The flood pulse concept: New aspects, INTRODUCTION plain, where production and degradation of organic matter also takes place. Rivers and floodplain wetlands are among the most threatened ecosystems. For example, 77 percent These characteristics are reflected for lakes in of the water discharge of the 139 largest river systems the “Seentypenlehre” (Lake typology), elaborated by in North America and Europe is affected by fragmen- Thienemann and Naumann between 1915 and 1935 tation of the river channels by dams and river regula- (e.g. -

Deposition Patterns and Rates of Mining-Contaminated Sediment Within a Sedimentation Basin System, S.E
BearWorks Institutional Repository MSU Graduate Theses Spring 2017 Deposition Patterns and Rates of Mining- Contaminated Sediment within a Sedimentation Basin System, S.E. Missouri Joshua Carl Voss Missouri State University - Springfield, [email protected] Follow this and additional works at: http://bearworks.missouristate.edu/theses Part of the Environmental Indicators and Impact Assessment Commons, Environmental Monitoring Commons, and the Hydrology Commons Recommended Citation Voss, Joshua Carl, "Deposition Patterns and Rates of Mining-Contaminated Sediment within a Sedimentation Basin System, S.E. Missouri" (2017). MSU Graduate Theses. 3074. http://bearworks.missouristate.edu/theses/3074 This article or document was made available through BearWorks, the institutional repository of Missouri State University. The orkw contained in it may be protected by copyright and require permission of the copyright holder for reuse or redistribution. For more information, please contact [email protected]. DEPOSITION PATTERNS AND RATES OF MINING-CONTAMINATED SEDIMENT WITHIN A SEDIMENTATION BASIN SYSTEM, BIG RIVER, S.E. MISSOURI A Masters Thesis Presented to The Graduate College of Missouri State University ATE In Partial Fulfillment Of the Requirements for the Degree Master of Science, Geospatial Sciences in Geography, Geology, and Planning By Josh C. Voss May 2017 Copyright 2017 by Joshua Carl Voss ii DEPOSITION PATTERNS AND RATES OF MINING-CONTAMINATED SEDIMENT WITHIN A SEDIMENTATION BASIN SYSTEM, BIG RIVER, S.E. MISSOURI Geography, Geology, and Planning Missouri State University, May 2017 Master of Science Josh C. Voss ABSTRACT Flooding events exert a dominant control over the deposition and formation of floodplains. The rate at which floodplains form depends on flood magnitude, frequency, and duration, and associated sediment transport capacity and supply. -

Ecotone Properties and Influences on Fish Distributions Along Habitat Gradients of Complex Aquatic Systems
ECOTONE PROPERTIES AND INFLUENCES ON FISH DISTRIBUTIONS ALONG HABITAT GRADIENTS OF COMPLEX AQUATIC SYSTEMS A Dissertation Presented to the Faculty of the Graduate School of Cornell University In Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy by Nuanchan Singkran May 2007 © 2007 Nuanchan Singkran ECOTONE PROPERTIES AND INFLUENCES ON FISH DISTRIBUTIONS ALONG HABITAT GRADIENTS OF COMPLEX AQUATIC SYSTEMS Nuanchan Singkran, Ph. D. Cornell University 2007 Ecotone properties (formation and function) were studied in complex aquatic systems in New York State. Ecotone formations were detected on two embayment- stream gradients associated with Lake Ontario during June–August 2002, using abrupt changes in habitat variables and fish species compositions. The study was repeated at a finer scale along the second gradient during June–August 2004. Abrupt changes in the habitat variables (water depth, current velocity, substrates, and covers) and peak species turnover rate showed strong congruence at the same location on one gradient. The repeated study on the second gradient in the summer of 2004 confirmed the same ecotone orientation as that detected in the summer of 2002 and revealed the ecotone width covering the lentic-lotic transitions. The ecotone on the second gradient acted as a hard barrier for most of the fish species. Ecotone properties were determined along the Hudson River estuary gradient during 1974–2001 using the same methods employed in the freshwater system. The Hudson ecotones showed both changes in location and structural formation over time. Influences of tide, freshwater flow, salinity, dissolved oxygen, and water temperature tended to govern ecotone properties. One ecotone detected in the lower-middle gradient portion appeared to be the optimal zone for fish assemblages, but the other ecotones acted as barriers for most fish species. -

Lesson 4: Sediment Deposition and River Structures
LESSON 4: SEDIMENT DEPOSITION AND RIVER STRUCTURES ESSENTIAL QUESTION: What combination of factors both natural and manmade is necessary for healthy river restoration and how does this enhance the sustainability of natural and human communities? GUIDING QUESTION: As rivers age and slow they deposit sediment and form sediment structures, how are sediments and sediment structures important to the river ecosystem? OVERVIEW: The focus of this lesson is the deposition and erosional effects of slow-moving water in low gradient areas. These “mature rivers” with decreasing gradient result in the settling and deposition of sediments and the formation sediment structures. The river’s fast-flowing zone, the thalweg, causes erosion of the river banks forming cliffs called cut-banks. On slower inside turns, sediment is deposited as point-bars. Where the gradient is particularly level, the river will branch into many separate channels that weave in and out, leaving gravel bar islands. Where two meanders meet, the river will straighten, leaving oxbow lakes in the former meander bends. TIME: One class period MATERIALS: . Lesson 4- Sediment Deposition and River Structures.pptx . Lesson 4a- Sediment Deposition and River Structures.pdf . StreamTable.pptx . StreamTable.pdf . Mass Wasting and Flash Floods.pptx . Mass Wasting and Flash Floods.pdf . Stream Table . Sand . Reflection Journal Pages (printable handout) . Vocabulary Notes (printable handout) PROCEDURE: 1. Review Essential Question and introduce Guiding Question. 2. Hand out first Reflection Journal page and have students take a minute to consider and respond to the questions then discuss responses and questions generated. 3. Handout and go over the Vocabulary Notes. Students will define the vocabulary words as they watch the PowerPoint Lesson. -

Seasonal Flooding Affects Habitat and Landscape Dynamics of a Gravel
Seasonal flooding affects habitat and landscape dynamics of a gravel-bed river floodplain Katelyn P. Driscoll1,2,5 and F. Richard Hauer1,3,4,6 1Systems Ecology Graduate Program, University of Montana, Missoula, Montana 59812 USA 2Rocky Mountain Research Station, Albuquerque, New Mexico 87102 USA 3Flathead Lake Biological Station, University of Montana, Polson, Montana 59806 USA 4Montana Institute on Ecosystems, University of Montana, Missoula, Montana 59812 USA Abstract: Floodplains are comprised of aquatic and terrestrial habitats that are reshaped frequently by hydrologic processes that operate at multiple spatial and temporal scales. It is well established that hydrologic and geomorphic dynamics are the primary drivers of habitat change in river floodplains over extended time periods. However, the effect of fluctuating discharge on floodplain habitat structure during seasonal flooding is less well understood. We collected ultra-high resolution digital multispectral imagery of a gravel-bed river floodplain in western Montana on 6 dates during a typical seasonal flood pulse and used it to quantify changes in habitat abundance and diversity as- sociated with annual flooding. We observed significant changes in areal abundance of many habitat types, such as riffles, runs, shallow shorelines, and overbank flow. However, the relative abundance of some habitats, such as back- waters, springbrooks, pools, and ponds, changed very little. We also examined habitat transition patterns through- out the flood pulse. Few habitat transitions occurred in the main channel, which was dominated by riffle and run habitat. In contrast, in the near-channel, scoured habitats of the floodplain were dominated by cobble bars at low flows but transitioned to isolated flood channels at moderate discharge. -

Probabilistic Extreme Flood Hydrographs That Use Paleoflood Data for Dam Safety Applications
Probabilistic Extreme Flood Hydrographs That Use PaleoFlood Data for Dam Safety Applications Dam Safety Office Report No. DSO-03-03 Department of the Interior Bureau of Reclamation June 2003 Contents Page Introduction................................................................................................................................... 1 1.1 Background..................................................................................................................................2 1.2 Objectives....................................................................................................................................4 1.3 Acknowledgements .....................................................................................................................4 Probabilistic Extreme Flood Hydrographs from Streamflow Sampling................................. 5 2.1 General Procedure .......................................................................................................................5 2.2 Example Applications................................................................................................................11 Probabilitic Extreme Flood Hydrographs Using Rainfall-Runoff Models............................ 18 3.1 General Procedure .....................................................................................................................19 3.2 Example Applications s.............................................................................................................20 Reservoir Routing -

Variable Hydrologic and Geomorphic Responses to Intentional Levee Breaches Along the Lower Cosumnes River, California
Received: 21 April 2016 Revised: 29 March 2017 Accepted: 30 March 2017 DOI: 10.1002/rra.3159 RESEARCH ARTICLE Not all breaks are equal: Variable hydrologic and geomorphic responses to intentional levee breaches along the lower Cosumnes River, California A. L. Nichols1 | J. H. Viers1,2 1 Center for Watershed Sciences, University of California, Davis, California, USA Abstract 2 School of Engineering, University of The transport of water and sediment from rivers to adjacent floodplains helps generate complex California, Merced, California, USA floodplain, wetland, and riparian ecosystems. However, riverside levees restrict lateral connectiv- Correspondence ity of water and sediment during flood pulses, making the re‐introduction of floodplain hydrogeo- A. L. Nichols, Center for Watershed Sciences, morphic processes through intentional levee breaching and removal an emerging floodplain University of California, Davis, California, USA. restoration practice. Repeated topographic observations from levee breach sites along the lower Email: [email protected] Cosumnes River (USA) indicated that breach architecture influences floodplain and channel hydrogeomorphic processes. Where narrow breaches (<75 m) open onto graded floodplains, Funding information California Department of Fish and Wildlife archetypal crevasse splays developed along a single dominant flowpath, with floodplain erosion (CDFW) Ecosystem Restoration Program in near‐bank areas and lobate splay deposition in distal floodplain regions. Narrow breaches (ERP), Grant/Award Number: E1120001; The opening into excavated floodplain channels promoted both transverse advection and turbulent Nature Concervancy (TNC); Consumnes River Preserve diffusion of sediment into the floodplain channel, facilitating near‐bank deposition and potential breach closure. Wide breaches (>250 m) enabled multiple modes of water and sediment transport onto graded floodplains. -

Stormwater Design Manual Chapter Three
CHAPTER 3 HYDROLOGY Chapter 3 HYDROLOGY Table of Contents 3.1 INTRODUCTION .................................................................................................... 2 3.2 HYDROLOGIC DESIGN POLICIES........................................................................ 2 3.2.1 FULLY DEVELO PED CONDITIONS......................................................................... 3 3.2.2 DRAINAG E AREA ............................................................................................... 4 3.2.3 RAINFALL DATA AND INTENSITY .......................................................................... 4 3.3 TIME OF CONCENTRATION ................................................................................. 4 3.3.1 SCS METHOD................................................................................................... 5 3.3.1.1 Lag Time .................................................................................................. 5 3.3.1.2 Travel Time .............................................................................................. 5 3.3.1.3 Sheet Flow ............................................................................................... 6 3.3.1.4 Shallow Concentrated Flow....................................................................... 7 3.3.1.5 Channelized Flow ..................................................................................... 7 3.3.2 KIRPICH EQUATION ........................................................................................... 8 3.4 RATIONAL METHOD............................................................................................ -

Biogeochemical and Metabolic Responses to the Flood Pulse in a Semi-Arid Floodplain
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by DigitalCommons@USU 1 Running Head: Semi-arid floodplain response to flood pulse 2 3 4 5 6 Biogeochemical and Metabolic Responses 7 to the Flood Pulse in a Semi-Arid Floodplain 8 9 10 11 with 7 Figures and 3 Tables 12 13 14 15 H. M. Valett1, M.A. Baker2, J.A. Morrice3, C.S. Crawford, 16 M.C. Molles, Jr., C.N. Dahm, D.L. Moyer4, J.R. Thibault, and Lisa M. Ellis 17 18 19 20 21 22 Department of Biology 23 University of New Mexico 24 Albuquerque, New Mexico 87131 USA 25 26 27 28 29 30 31 present addresses: 32 33 1Department of Biology 2Department of Biology 3U.S. EPA 34 Virginia Tech Utah State University Mid-Continent Ecology Division 35 Blacksburg, Virginia 24061 USA Logan, Utah 84322 USA Duluth, Minnesota 55804 USA 36 540-231-2065, 540-231-9307 fax 37 [email protected] 38 4Water Resources Division 39 United States Geological Survey 40 Richmond, Virginia 23228 USA 41 1 1 Abstract: Flood pulse inundation of riparian forests alters rates of nutrient retention and 2 organic matter processing in the aquatic ecosystems formed in the forest interior. Along the 3 Middle Rio Grande (New Mexico, USA), impoundment and levee construction have created 4 riparian forests that differ in their inter-flood intervals (IFIs) because some floodplains are 5 still regularly inundated by the flood pulse (i.e., connected), while other floodplains remain 6 isolated from flooding (i.e., disconnected). -

Effects of Beaver Dams on Benthic Macroinvertebrates
Effects ofbeaver dams onbenthic macroinvertebrates Andreas Johansson Degree project inbiology, Master ofscience (2years), 2014 Examensarbete ibiologi 45 hp tillmasterexamen, 2014 Biology Education Centre, Uppsala University, and Department ofAquatic Sciences and Assessment, SLU Supervisor: Frauke Ecke External opponent: Peter Halvarsson ABSTRACT In the 1870's the beaver (Castor fiber), population in Sweden had been exterminated. The beaver was reintroduced to Sweden from the Norwegian population between 1922 and 1939. Today the population has recovered and it is estimated that the population of C. fiber in all of Europe today ranges around 639,000 individuals. The main aim with this study was to investigate if there was any difference in species diversity between sites located upstream and downstream of beaver ponds. I found no significant difference in species diversity between these sites and the geographical location of the streams did not affect the species diversity. This means that in future studies it is possible to consider all streams to be replicates despite of geographical location. The pond age and size did on the other hand affect the species diversity. Young ponds had a significantly higher diversity compared to medium-aged ponds. Small ponds had a significantly higher diversity compared to medium-sized and large ponds. The upstream and downstream reaches did not differ in terms of CPOM amount but some water chemistry variables did differ between them. For the functional feeding groups I only found a difference between the sites for predators, which were more abundant downstream of the ponds. SAMMANFATTNING Under 1870-talet utrotades den svenska populationen av bäver (Castor fiber). -

Classifying Rivers - Three Stages of River Development
Classifying Rivers - Three Stages of River Development River Characteristics - Sediment Transport - River Velocity - Terminology The illustrations below represent the 3 general classifications into which rivers are placed according to specific characteristics. These categories are: Youthful, Mature and Old Age. A Rejuvenated River, one with a gradient that is raised by the earth's movement, can be an old age river that returns to a Youthful State, and which repeats the cycle of stages once again. A brief overview of each stage of river development begins after the images. A list of pertinent vocabulary appears at the bottom of this document. You may wish to consult it so that you will be aware of terminology used in the descriptive text that follows. Characteristics found in the 3 Stages of River Development: L. Immoor 2006 Geoteach.com 1 Youthful River: Perhaps the most dynamic of all rivers is a Youthful River. Rafters seeking an exciting ride will surely gravitate towards a young river for their recreational thrills. Characteristically youthful rivers are found at higher elevations, in mountainous areas, where the slope of the land is steeper. Water that flows over such a landscape will flow very fast. Youthful rivers can be a tributary of a larger and older river, hundreds of miles away and, in fact, they may be close to the headwaters (the beginning) of that larger river. Upon observation of a Youthful River, here is what one might see: 1. The river flowing down a steep gradient (slope). 2. The channel is deeper than it is wide and V-shaped due to downcutting rather than lateral (side-to-side) erosion. -
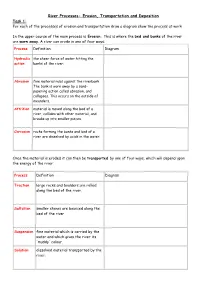
River Processes- Erosion, Transportation and Deposition Task 1: for Each of the Processes of Erosion and Transportation Draw a Diagram Show the Process at Work
River Processes- Erosion, Transportation and Deposition Task 1: For each of the processes of erosion and transportation draw a diagram show the process at work In the upper course of the main process is Erosion. This is where the bed and banks of the river are worn away. A river can erode in one of four ways: Process Definition Diagram Hydraulic the sheer force of water hitting the action banks of the river: Abrasion fine material rubs against the riverbank The bank is worn away by a sand- papering action called abrasion, and collapses. This occurs on the outside of meanders. Attrition material is moved along the bed of a river, collides with other material, and breaks up into smaller pieces. Corrosion rocks forming the banks and bed of a river are dissolved by acids in the water. Once the material is eroded it can then be transported by one of four ways, which will depend upon the energy of the river: Process Definition Diagram Traction large rocks and boulders are rolled along the bed of the river. Saltation smaller stones are bounced along the bed of the river Suspension fine material which is carried by the water and which gives the river its 'muddy' colour. Solution dissolved material transported by the river. In the middle and lower course, the land is much flatter, this means that the river is flowing more slowly and has much less energy. The river starts to deposit (drop) the material that it has been carry Deposition Challenge: Add labels onto the diagram to show where all of the processes could be happening in the river channel.