Haplopappus Crispus and H. Zionis (Asteradeae): New Species from Utah
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Notes on Variation and Geography in Rayjacksonia Phyllocephala (Asteraceae: Astereae)
Nesom, G.L., D.J. Rosen, and S.K. Lawrence. 2013. Notes on variation and geography in Rayjacksonia phyllocephala (Asteraceae: Astereae). Phytoneuron 2013-53: 1–15. Published 12 August 2013. ISSN 2153 733X NOTES ON VARIATION AND GEOGRAPHY IN RAYJACKSONIA PHYLLOCEPHALA (ASTERACEAE: ASTEREAE) GUY L. NESOM 2925 Hartwood Drive Fort Worth, Texas 76109 [email protected] DAVID J. ROSEN Department of Biology Lee College Baytown, Texas 77522-0818 [email protected] SHIRON K. LAWRENCE Department of Biology Lee College Baytown, Texas 77522-0818 ABSTRACT Inflorescences of Rayjacksonia phyllocephala in the disjunct Florida population system are characterized by heads on peduncles with leaves mostly reduced to linear bracts; heads in inflorescences of the Mexico-Texas-Louisiana system are immediately subtended by relatively unreduced leaves. The difference is consistent and justifies recognition of the Florida system as R. phyllocephala var. megacephala (Nash) D.B. Ward. Scattered waifs between the two systems are identified here as one or the other variety, directly implying their area of origin. In the eastern range of var. phyllocephala , at least from Brazoria County, Texas, eastward about 300 miles to central Louisiana, leaf margins vary from entire to deeply toothed-spinulose. In contrast, margins are invariably toothed-spinulose in var. megacephala as well as in the rest of southeastern Texas (from Brazoria County southwest) into Tamaulipas, Mexico. In some of the populations with variable leaf margins, 70-95% of the individuals have entire to mostly entire margins. KEY WORDS : Rayjacksonia , morphological variation, leaf margins, disjunction, waifs Rayjacksonia phyllocephala (DC.) Hartman & Lane (Gulf Coast camphor-daisy) is an abundant and conspicuous species of the shore vegetation around the Gulf of Mexico. -

Phytologia an International Journal to Expedite
PHYTOLOGIA An intern ational jou rnal to ex edite la n t s stematic b to eo ra bi l p p y , p y g g p ca and ecological pu blication 1 S e ember 1 1 Vo l. 7 pt 99 CONTENTS TRE M N w ll n n n t r c l fl r . A ne CUA CASAS . c o o o o o o , J , is e a e us tes e pi a a XX species of Humiriastru m 1 65 R T F w f S . rm l rr t n f t c th in the th rn OSS . o co c o o o c t o , , a e i spe i i epi e s s u e . N N H N A eratina nd w f B artlettina ROB S O . o t on a ne c o / I , , es g a spe ies (Eupato rieae : Asteraceae) 1 7 1 /R )BIN N H N w w f C SO . e c and ne comb n on o Crit oniinae from , , spe ies i ati s Meso ame ri ca (Eupato rieae : Asteraceae) 1 76 ’ R B N N H Tw w f Fleiscbman m a fr m M r O S O . o ne c o o m c / I , , spe ies o es a e i a (Eupatorieae : As t eraceae) 1 8 1 /ROBIN N Tw w M le a M SO H. o ne c o f i an i in m r c m , , spe ies esoa e i a (Eupatorieae : As te raceae) 1 84 E AL N D L f f a a r i S C O A F . -
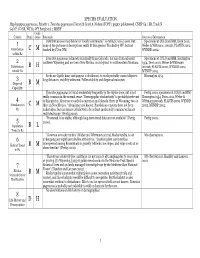
1 C M 2 B H 3 B M 4 C M 5 B L 6 B M 7 B M 8
SPECIES EVALUATION Haplopappus pygmaeus, Priority 1. Tonestus pygmaeus (Torrey & Gray) A. Nelson (TOPY). pygmy goldenweed. CNHP G4 / SR, Track N G4 N?. CO SR, WY S1. WY Peripheral 2 MBNF Confi- Criteria Rank dence Rationale Sources of Information Distribution (see map below) is “nearly continuous,” so rating C was chosen, but Specimens at COLO and RM, Dorn 2001, 1 none of the pictures or descriptions really fit this species. Tracked by WY, but not Weber & Wittmann 2001ab, PLANTS 2002, Distribution C M tracked by CO or NM. WYNDD 2002. within R2 Tonestus pygmaeus is known principally from Colorado, but also from adjacent Specimens at COLO and RM, Harrington 2 southern Wyoming and northern New Mexico, and disjunct in southwestern Montana. 1954, Dorn 2001, Weber & Wittmann Distribution B H 2001ab, PLANTS 2002, WYNDD 2002, outside R2 MTNHP 2002. Seeds are lightly hairy and pappus is deciduous, so seeds probably cannot disperse Harrington 1954. 3 long distances; viability unknown. Pollen viability and dispersal unknown. Dispersal B M Capability Tonestus pygmaeus is found moderately frequently in the Alpine zone, but is not Fertig 2000, specimens at COLO and RM, really common in the normal sense. “Demographic stochasticity” is probably irrelevant Harrington 1954, Dorn 2001, Weber & 4 to this species. About 50 recorded occurrences in Colorado, three in Wyoming, two or Wittmann 2001ab, PLANTS 2002, WYNDD Abundance in C M three in New Mexico. “Abundance not known. Population censuses have not been 2002, MTNHP 2002. R2 undertaken, but the species is believed to be at least moderately common within its restricted range” (Fertig 2000). -

Reclassification of North American Haplopappus (Compositae: Astereae) Completed: Rayjacksonia Gen
AmericanJournal of Botany 83(3): 356-370. 1996. RECLASSIFICATION OF NORTH AMERICAN HAPLOPAPPUS (COMPOSITAE: ASTEREAE) COMPLETED: RAYJACKSONIA GEN. NOV.1 MEREDITH A. LANE2 AND RONALD L. HARTMAN R. L. McGregor Herbarium(University of Kansas NaturalHistory Museum Division of Botany) and Departmentof Botany,University of Kansas, Lawrence, Kansas 66047-3729; and Rocky MountainHerbarium, Department of Botany,University of Wyoming,Laramie, Wyoming82071-3165 Rayjacksonia R. L. Hartman& M. A. Lane, gen. nov. (Compositae: Astereae), is named to accommodate the "phyllo- cephalus complex," formerlyof Haplopappus Cass. sect. Blepharodon DC. The new combinationsare R. phyllocephalus (DC.) R. L. Hartman& M. A. Lane, R. annua (Rydb.) R. L. Hartman& M. A. Lane, and R. aurea (A. Gray) R. L. Hartman & M. A. Lane. This transfercompletes the reclassificationof the North American species of Haplopappus sensu Hall, leaving that genus exclusively South American.Rayjacksonia has a base chromosomenumber of x = 6. Furthermore,it shares abruptlyampliate disk corollas, deltatedisk style-branchappendages, and corolla epidermalcell type,among other features,with Grindelia, Isocoma, Olivaea, Prionopsis, Stephanodoria, and Xanthocephalum.Phylogenetic analyses of morphologicaland chloroplastDNA restrictionsite data, taken together,demonstrate that these genera are closely related but distinct. Key words: Astereae; Asteraceae; Compositae; Haplopappus; Rayjacksonia. During the past seven decades, taxonomic application lopappus sensu Hall (1928) are reclassifiedand are cur- -

Florida Golden Aster Chrysopsis (=Heterotheca) Floridana Small
Florida Golden Aster Chrysopsis (=Heterotheca) floridana Small he Florida golden aster is a perennial herb in the Federal Status: Endangered (May 16, 1986) aster family with a distribution limited to a few Critical Habitat: None Designated Tcounties in west-central Florida. The Florida golden Florida Status: Endangered aster occurs in sand pine and oak scrub or in disturbed areas at the edges of scrub. Recovery Plan Status: Contribution (May 1999) This account represents South Floridas contribution to Geographic Coverage: South Florida the existing recovery plan for the Florida golden aster (FWS 1988). Figure 1. County distribution of the Florida golden aster. Description Chrysopsis floridana is a perennial herb with stems that are woody toward the base and non-woody above. The plants have basal rosettes (clusters of leaves at ground level) with leaves 4 to 10 cm long, 1.5 to 2.0 cm wide; the leaves of the rosette are densely short-wooly pubescent. The stem leaves are nearly the same size from the top to the bottom of the stem; they are obovate-elliptic, slightly clasping the stem, entire, and densely short-wooly pubescent. The flower heads are grouped into a more or less flat-topped cluster of 1 to 25 heads at the top of the stem. Each head is slightly over 2.5 cm in diameter. Both the central disc and the rays are golden yellow. C. floridana is distinguished from other members of its genus by its perennial habit, the woodiness of its stems, the wooliness and the shape of the stem and the leaves, and the way the flower heads are arranged in a flat-topped cluster (Semple 1981, Wunderlin et al. -

Jessica's Aster
Jessica’s Aster (Aster jessicae) Population Monitoring: Third-year Results Idaho Conservation Data Center Idaho Department of Fish and Game PO Box 25 Juanita Lichthardt Boise, Idaho Karen Gray 83707 2005 Report prepared for the U.S. Fish and Wildlife Service ABSTRACT Jessica’s aster (Aster jessicae) is a tall, rhizomatous aster endemic to the Palouse region of southeastern Washington and adjacent Idaho. Its habitat has been severely reduced by conversion of this region to intensive agricultural uses. It is restricted almost entirely to private lands, and because of this, little was known of its distribution and abundance until a 1991 status survey was conducted. Five permanent monitoring plots were established in 2001, to track and compare populations occupying contrasting sites. Monitoring sites differ in amount of edge, degree of isolation from other populations, habitat extent, and amount of forest cover. This report summarizes three consecutive years of monitoring data. i TABLE OF CONTENTS ABSTRACT ......................................................................................................................................i TABLE OF CONTENTS ................................................................................................................ ii LIST OF TABLES ......................................................................................................................... iii LIST OF FIGURES....................................................................................................................... -

Pityopsis) Graminifolia (Asteraceae: Astereae
Semple, J.C. and F. Jabbour. 2019. Type specimens of Inula (Pityopsis) graminifolia (Asteraceae: Astereae). Phytoneuron 2019-22. 1-9. Published 25 April 2019. ISSN 2153 733X TYPE SPECIMENS OF INULA (PITYOPSIS) GRAMINIFOLIA (ASTERACEAE: ASTEREAE) JOHN C. SEMPLE Department of Biology, University of Waterloo Waterloo, Ontario Canada N2L 3G1 [email protected] and FLORIAN JABBOUR Institut de Systématique, Évolution, Biodiversité (ISYEB) Muséum national d’Histoire naturelle, CNRS, Sorbonne Université, EPHE, Université des Antilles, 57 rue Cuvier, CP39, 75005 Paris, France ABSTRACT Recent proposed changes to the nomenclature of the goldenaster genus Pityopsis disagreed significantly on the application of the name Inula graminifolia, the basionym of Pityopsis graminifolia. In order to clarify the problem, the three known collections of Inula graminifolia in the Herbarium of the Muséum national d’Histoire naturelle, Paris (P) collected by André Michaux were examined. The holotype collection has small heads with many stipitate glands on the distal portion of the phyllaries and obvious long hairs on the peduncles. This is also the case for the isotype collection. A third collection by Michaux is mixed with a shoot with larger heads and shoots with smaller heads and all with phyllaries with few or no stipitate glands. Thus, the third specimen is not part of the same taxon as the holotype and isotype. The Pityopsis graminifolia (Michx.) Nutt. complex includes goldenasters native to the southeastern USA, southern Mexico, Guatemala, Belize, western Honduras, and the Bahamas. The complex includes either a single species with multiple varieties (Semple & Bowers 1985; Semple 2006) or the complex might be divided into multiple species with more narrowly defined morphological and ecological limits. -

FLORATECH New Crop Report
FLORATECH New Crop Report presents: Machaeranthera gracilis Slender Goldenweed Taxonomy Machaeranthera gracilis (Nutt.) Shinners Translated: sword-like anthers; graceful. A.k.a.: Dieteria gracilis Haplopappus gracilis Haplopappus ravenii Sideranthus gracilis And newly named Xanthisma gracile WITHIN ASTERACEAE GEOGRAPHIC DISTRIBUTION Native to CA, NV, UT, AZ, CO, NM, TX, ME, NY according to USDA NRCS No reports of naturalizing Climactic Conditions5: Sandy or rocky places to 5000’ Habitats: creosote bush scrub, joshua tree woodland, east Mojave Desert Taxonomic Description7 Annual with erect stems, 3.25cm, leafy, branched at or above base, bristly throughout. Leaves: 1-3cm long, 3-7mm wide; lower oblanceolate, elliptic, or oblong in outline, 1-2 pinnately lobed; upper linear, reduced, lobes and teeth bristle-tipped. Inflorescence: heads radiate, solitary or in cymes; involucre 6-7mm, 7-12mm wide, hemispheric; phyllaries (bracts) in rows of 4-6, linear-lanceolate, bristle-tipped, hairy. Flowering time often from April-June. Ray flower: 16-18; ligules 7-12mm, yellow. Disk flower: 44-65; corollas 4.5-5.5mm, yellow. Taxonomic Description cont. Machaeranthera spp. tend to have several short rhizomes arising from the caudex (woody base of a perennial).1 Rhizomes may or may not function in vegetative reproduction.1 Biseasonal (germinates in winter, but flowers and sets seed in summer.2 Seed set greatly varied from season to season and shows significant responses to microhabitat differentiation.2 Use by indigenous people The Ramah Navaho used M. gracilis3: In a cold infusion used as lotion for pimples, boils and sores. In a cold, compound infusion of plant used as an eyewash. -

Asters of Yesteryear (Updated April 2018)
Asters of Yesteryear (Updated April 2018) About this Update: The document was originally posted in a shorter version, to accompany the brief article "Where Have all our Asters Gone?" in the Fall 2017 issue of Sego Lily. In that version it consisted simply of photos of a number of plants that had at some time been included in Aster but that no longer are, as per Flora of North America. In this version I have added names to the photos to indicate how they have changed since their original publication: Date and original name as published (Basionym) IF name used in Intermountain Flora (1994) UF name used in A Utah Flora (1983-2016) FNA name used in Flora of North America (2006) I have also added tables to show the renaming of two groups of species in the Astereae tribe as organized in Intermountain Flora. Color coding shows how splitting of the major genera largely follows fault lines already in place No color Renamed Bright Green Conserved Various Natural groupings $ Plant not in Utah It is noteworthy how few species retain the names used in 1994, but also how the renaming often follows patterns already observed. Asters of Yesteryear (Updated April 2018) Here are larger photos (16 inches wide or tall at normal screen resolution of 72 dpi) of the plants shown in Sego Lily of Fall 2017, arranged by date of original publication. None of them (except Aster amellus on this page) are now regarded as true asters – but they all were at one stage in their history. Now all are in different genera, most of them using names that were published over 100 years ago. -

Report on the Conservation Status of Haplopappus Radiatus, in Idaho
REPORT ON THE CONSERVATION STATUS OF HAPLOPAPPUS RADIATUS, IN IDAHO by Michael Mancuso and Robert K. Moseley Conservation Data Center Nongame and Endangered Wildlife Program June, 1993 Idaho Department of Fish and Game 600 South Walnut, P.O. Box 25 Boise, Idaho 83707 Jerry M. Conley, Director Status Survey Report prepared for Idaho Department of Parks and Recreation through Section 6 funding from U.S. Fish and Wildlife Service, Region 1 REPORT ON THE CONSERVATION STATUS OF i HAPLOPAPPUS RADIATUS, IN IDAHO Taxon Name: Haplopappus radiatus (Nutt.) Cronq. Common Name: Snake River goldenweed Family: Asteraceae (Compositae) States Where Taxon Occurs: U.S.A.; Idaho, Oregon Current Federal Status: Category 1 Candidate Recommended Federal Status: Threatened Authors of Report: Michael Mancuso and Robert K. Moseley Original Date of Report: June 18, 1993 Date of Most Recent Revision: N/A Individual to Whom Further Information and Comments Should be Sent: Robert K. Moseley Conservation Data Center Idaho Dept. Fish and Game P.O. Box 25 Boise, ID 83707 ii ABSTRACT Snake River goldenweed (Haplopappus radiatus) has been recognized as a possible conservation concern for at least twenty years. Until 1991, field inventories in Idaho were limited. In 1991, the Idaho Conservation Data Center completed a field investigation for Snake River goldenweed on the Payette National Forest. Through funding provided by the U.S. Fish and Wildlife Service, additional fieldwork was completed throughout the rest of the species' range in Idaho, during 1992. Survey work has also been done in Oregon. Besides field inventories, research biologists associated with the Conservation Biology Program, Oregon Department of Agriculture, have done cytological and pollination investigations, and presently are monitoring several populations in Oregon. -

Redalyc.Biomass, Resin and Essential Oil Content and Their Variability In
Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas ISSN: 0717-7917 [email protected] Universidad de Santiago de Chile Chile GONZÁLEZ, Benita; VOGEL, Hermine; RAZMILIC, Iván; MARTÍN, José San; DOLL, Ursula Biomass, resin and essential oil content and their variability in natural populations of the Chilean crude drug "bailahuén" (Haplopappus spp.) Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas, vol. 11, núm. 1, 2012, pp. 66-73 Universidad de Santiago de Chile Santiago, Chile Available in: http://www.redalyc.org/articulo.oa?id=85622229007 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative © 2012 Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas 11 (1): 66 - 73 ISSN 0717 7917 www.blacpma.usach.cl Artículo Original | Original Article Biomass, resin and essential oil content and their variability in natural populations of the Chilean crude drug “bailahuén” (Haplopappus spp.) [Biomasa, contenido de resina y aceite esencial y su variabilidad en poblaciones naturales de las especies de droga cruda chilena “bailahuén” (Haplopappus spp.)] Benita GONZÁLEZ1, Hermine VOGEL1, Iván RAZMILIC2, José San MARTÍN3 & Ursula DOLL4 1 Facultad de Ciencias Agrarias, Universidad de Talca, Talca, Chile. 2 Instituto de Química de Recursos Naturales, Universidad de Talca, Talca, Chile. 3 Instituto de Biología Vegetal y Biotecnología, Universidad de Talca, Talca, Chile. 4 Facultad de Ciencias Forestales, Universidad de Talca, Talca, Chile. Contactos | Contacts: Benita GONZALEZ - E-mail address: [email protected] Abstract Bailahuén (Haplopappus rigidus, Haplopappus baylahuen, Haplopappus multifolius and Haplopappus taeda; Asteraceae) are medicinal shrubs native to the Andes Mountains of Chile widely used to treat hepatic ailments. -

Origin of the Rapa Endemic Genus Apostates: Revisiting Major Disjunctions and Evolutionary Conservatism in the Bahia Alliance (Compositae: Bahieae) Bruce G
Baldwin & Wood • Systematics and biogeography of the Bahia alliance TAXON 65 (5) • October 2016: 1064–1080 Origin of the Rapa endemic genus Apostates: Revisiting major disjunctions and evolutionary conservatism in the Bahia alliance (Compositae: Bahieae) Bruce G. Baldwin1 & Kenneth R. Wood2 1 Jepson Herbarium and Department of Integrative Biology, University of California, Berkeley, California 94720, U.S.A. 2 National Tropical Botanical Garden, Kalaheo, Hawaii 96741, U.S.A. Author for correspondence: Bruce G. Baldwin, [email protected] ORCID BGB, http://orcid.org/0000-0002-0028-2242; KRW, http://orcid.org/0000-0001-6446-1154 DOI http://dx.doi.org/10.12705/655.8 Abstract Molecular phylogenetic analyses of nuclear and chloroplast DNA sequences indicate that the rediscovered Apostates, known only from Rapa in the Austral Islands of southeastern Polynesia, represents an example of extreme long-distance dispersal (> 6500 km) from southwestern North America and one of at least four disjunctions of comparable magnitude in the primarily New World Bahia alliance (tribe Bahieae). Each of the disjunctions appears to have resulted from north-to-south dispersal since the mid-Miocene; three are associated with such marked morphological and ecological change that some of the southern taxa (including Apostates) have been treated in distinct genera of uncertain relationship. Phyllotaxy within the Bahia alliance, however, evidently has been even more conservative evolutionarily than reflected by previous taxonomies, with alternate-leaved and opposite-leaved clades in Bahia sensu Ellison each encompassing representatives of other genera that share the same leaf arrangements. A revised taxonomic treatment of the Bahia alliance is proposed to recognize morphologically distinctive, monophyletic genera, including the critically endangered Apostates.