Growth in Four Populations of Leporinus Friderici
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Identification Des Contraintes Socio-Économiques À L
REGION GUYANE CENTRE DE CAYENNE PROGRAMME INTEGRE D'AMENAGEMENT RURAL (PIAR) IDENTIFICAnON DES CONTRAINTES SOCIO-ECONOMIQUES A L'INSTALLATION D'AGRICULTEURS SUR LE PERIMETRE DE ROCOUCOUA (IRACOUBO) C.LEBERRE AVRIL 1990 Convention entre le Conseil Régional de la Guyane et l'Institut Français de Recherche Scientifique pour le Développement en Coopération (ORSTOM) AVANT-PROPOS Dans le cadre de la politique de développement agricole mise en oeuvre par le Conseil Régional de la Guyane, le Centre ORSTOM de Cayenne est sollicité pour effectuer plusieurs études agro- pédologiques et socio-économiques. L'objet de ces études est défini par les termes d'une convention, signée le 9 mars 1989, entre la Région Guyane et 1 'ORSTOM, "relative à la mise en place des Programmes Intégrés d'Aménagement Rural (PIAR}". Celle-ci prévoyait sur la zone de Rocoucoua à Iracoubo "des travaux de recherche relatifs à la structuration de l'espace rural, liés à l'installation des agriculteurs", à savoir: - "l'évaluation des possibilités d' extension du périmètre vers le Sud-Ouest dans la limite des zones accessibles depuis la piste; - le recensement des principales contraintes rencontrées par les agriculteurs". Dans ce document sont présentés les résultats des travaux socio-économiques, effectués à Iracoubo. Ceux-ci visent à déterminer les principales contraintes rencontrées par les agriculteurs "professionnels" sur le périmètre de Rocoucoua. Ces travaux se rattachent à une activité de recherche plus globale du Centre ORSTOM de Cayenne, concernant les stratégies et les politiques de développement des filières de production, animée par J. MICHOTTE, Directeur de Recherche. Il existe en Guyane une volonté de développer le secteur agricole. -

Proposition Pour Une Clé D'identification Des Variétés De
Colloques du Groupement des Anthropologistes de Langue Française (GALF) Proposition pour une clé d’identification des variétés de manioc chez différents groupes culturels en Guyane française A data record for describe the cassava varieties in a diversity of human cultural groups in French Guiana Sandrine Manusset E-mail: [email protected] Mots-clés: manioc, agro-biodiversité, classification locale, description morphologique, clé d’identification, Guyane française Key-words: cassava, varietal diversity, local classification, data record, French Guiana Résumé Cet article s’inscrit dans le développement des stratégies de conservation in situ de la biodiversité, dans le cadre duquel il convient simultanément de comprendre les dynamiques sociales (et biologiques) responsables de l’origine et du maintien de la diversité des ressources génétiques et d’élaborer des outils et des méthodes pour appuyer la mise en place et le suivi de programmes de gestion et de valorisation de la biodiversité. S’appuyant sur une connaissance approfondie de la pratique agricole de l’abattis (Manusset, 2004a, b et c) au sein delaquelle la question de l’agro-biodiversité du manioc, Manihot esculenta, s’intègre en Guyane, la présentation de l’étude du cas guyanais répond à ce courant de recherche en trois points. A partir des résultats de plusieurs inventaires des variétés de manioc, réalisés chez différents groupes culturels, une évaluation du niveau de la diversité variétale de cette plante cultivée est présentée en s’appuyant sur deux indicateurs: le nombre total de variétés recensées par groupe culturel ou par région (N) et le nombre moyen de variétés recensées par exploitant (NE). Ensuite, la pratique sociale de gestion de la diversité est abordée à travers la description minutieuse des processus locaux de dénomination et de sélection des variétés. -

How Can the Freshwater Outflow from the Amazon Increase the Salinity of the Guianan Shore?
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Open Archive Toulouse Archive Ouverte The Guianese paradox: How can the freshwater outflow from the Amazon increase the salinity of the Guianan shore? Luc Lambs *, Etienne Muller, F. Fromard EcoLab – Laboratoire d’e´cologie fonctionnelle, UMR 5245 (CNRS-UPS-INPT), 29 rue Jeanne Marvig, 31055 Toulouse cedex, France KEYWORDS Summary French Guiana is notable for the extent of its rain forests, which occupy 97% of French Guiana rivers; the country, and the influence of the Amazon along its shores. In fact, the shores and estu- Stable isotopes; aries support a mangrove forest typical of saline conditions. This paper reports the chem- Oxygen-18; ical characteristics, conductivity and salinity and the stable isotopes (oxygen and Deuterium; deuterium) of the rivers and shores between the Cayenne area and the border with Suri- Salinity nam. The results show a quite homogenous freshwater pool over the country. However, the low slope of the coast, a result of the wide mud banks deposited by the Amazonian plume, have turned the mouths of the smaller rivers to the northwest, creating large salty areas where mangroves grow several kilometers inland. Despite the large amount of Ama- zonian water, the Guianan coast exhibits high salinity. In fact, the freshwater itself remains far from the shore, following the north Brazilian current, while only the mud plume arrives at the coast, creating this paradox. Introduction lens. The interaction of Amazon water and seawater, in front of the Amazonian estuary, were studied during the The discharge from many large rivers, because of the lower Amaseds program between 1989 and 1991 (Karr and Show- density of freshwater relative to seawater, can be detected ers, 2002). -

Panorama # 3 Propriété Foncière Atlas Cartographique 2017
Réalisé dans le cadre de l'observatoire des sols et cartes communales). Ces A qui appartient la Guyane ? De quels foncier de l'AUDeG, cet atlas cartogra- croisements fournissent une lecture types de terrains l’Etat est-il proprié- phique se propose de faire un tour dynamique du foncier, mettant en taire ? Quels sont ceux qui appartien- d'horizon de la situation de la proprié- regard propriété des terrains et desti- nent aux collectivités ? Qu’en est-il à té foncière dans le temps et dans l'es- nation des sols. l’échelle des communes ? Quelle est la pace afin d’apporter des éléments L'atlas s'intéresse également à la pro- destination des terrains privés dans les permettant d'alimenter une réflexion priété au sein des 24 secteurs définis documents d’urbanisme ? Comment la globale sur ce sujet. dans le cadre de l’OIN (opération propriété a-t-elle évolué entre 2007 et Mettant en perspective propriété et d’intérêt national). 2017 ? planification, il explore les questions La comparaison des millésimes sur la Ce sont là autant d’interrogations de l’organisation de l’espace, à travers dernière décennie (2007-2017) permet auxquelles l’atlas cartographique de la le SAR (schéma d’aménagement régio- quant à elle de suivre les évolutions propriété foncière entend fournir des nal), mais aussi à travers les docu- temporelles du cadastre et de la pro- éléments de réponse. ments d’urbanisme communaux (plan priété sur les dix dernières années. locaux d’urbanisme, plan d’occupation OBSFONCIER > Panorama #2 - Septembre 2018 - Page 1 Observatoire foncier de la Guyane | AUDeG Page 2 - OBSFONCIER > Panorama #2 - Septembre 2018 Observatoire foncier de la Guyane | AUDeG Une connaissance de la propriété limitée ............................................................................................................ -

Phylogenetic Relationships Within the Speciose Family Characidae
Oliveira et al. BMC Evolutionary Biology 2011, 11:275 http://www.biomedcentral.com/1471-2148/11/275 RESEARCH ARTICLE Open Access Phylogenetic relationships within the speciose family Characidae (Teleostei: Ostariophysi: Characiformes) based on multilocus analysis and extensive ingroup sampling Claudio Oliveira1*, Gleisy S Avelino1, Kelly T Abe1, Tatiane C Mariguela1, Ricardo C Benine1, Guillermo Ortí2, Richard P Vari3 and Ricardo M Corrêa e Castro4 Abstract Background: With nearly 1,100 species, the fish family Characidae represents more than half of the species of Characiformes, and is a key component of Neotropical freshwater ecosystems. The composition, phylogeny, and classification of Characidae is currently uncertain, despite significant efforts based on analysis of morphological and molecular data. No consensus about the monophyly of this group or its position within the order Characiformes has been reached, challenged by the fact that many key studies to date have non-overlapping taxonomic representation and focus only on subsets of this diversity. Results: In the present study we propose a new definition of the family Characidae and a hypothesis of relationships for the Characiformes based on phylogenetic analysis of DNA sequences of two mitochondrial and three nuclear genes (4,680 base pairs). The sequences were obtained from 211 samples representing 166 genera distributed among all 18 recognized families in the order Characiformes, all 14 recognized subfamilies in the Characidae, plus 56 of the genera so far considered incertae sedis in the Characidae. The phylogeny obtained is robust, with most lineages significantly supported by posterior probabilities in Bayesian analysis, and high bootstrap values from maximum likelihood and parsimony analyses. -

ESPÉCIES NOME POPULAR FONTE Classe ACTINOPTERYGII Ordem
A NEXO 11: Lista de espécies de peixes coletadas no Ribeirão Claro (SP). 1- Referente à dissertação de mestrado de Alexandre Tadeu Barbosa dos Santos, em andamento. 2 - Referente ao trab alho de iniciação científica de André Teixeira da Silva, em andamento. OBS: Ambos os estudo s estão sendo realizados pelo Departamento de Zoologia – IB – UNESP – Rio Claro. NOME ESPÉCIES FONTE POPULAR Classe ACTINOPTERYGII Ordem CHARACIFORMES Família ANOSTOMIDAE CETRA (2003); SANTOS (dados não publicados)1; Leporinus octofasciatus piau SILVA (dados não publicados) 2 CETRA (2003); SANTOS (dados não publicados) 1; Schizodon nasutus ximborê, taguara SILVA (dados não publicados) 2 Família CHARACIDAE SANTOS (dados não publicados) 1; SILVA (dados Acestrorhynchus lacustris peixe-cachorro não publicados) 2 CETRA (2003); SANTOS (dados não publicados) 1; Astyanax altiparanae tambiu SILVA (dados não publicados) 2 Astyanax fasciatus lambari do rabo CETRA (2003); SANTOS (dados não publicados) 1 vermelho Astyanax scabripinis paranae lambari CETRA (2003); SANTOS (dados não publicados) 1 Cheirodon stenodon pequira SANTOS (dados não publicados) 1 CETRA (2003); SANTOS (dados não publicados) 1; Hyphessobrycon eques mato-grosso SILVA (dados não publicados) 2 Odontostilbe cf. sp. pequira SANTOS (dados não publicados) 1 Piabina argentea pequira SANTOS (dados não publicados) 1 Planautina sp. pequira SANTOS (dados não publicados) 1 CETRA (2003); SANTOS (dados não publicados) 1; Salminus hilarii tabarana SILVA (dados não publicados) 2 CETRA (2003); SANTOS (dados não publicados) 1; Serrapinus heterodon pequira SILVA (dados não publicados) 2 CETRA (2003); SANTOS (dados não publicados) 1; Serrapinus notomelas pequira SILVA (dados não publicados) 2 CETRA (2003); SANTOS (dados não publicados) 1; Serrasalmus spilopleura pirambeba SILVA (dados não publicados) 2 Família CRENICHIDAE Characidium cf. -
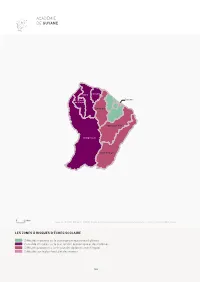
Académie De Guyane
ACADÉMIE DE GUYANE Mana Iracoubo St-Laurent- Cayenne du-Maroni Sinnamary Approuage-Kaw Maripasoula Saint-Georges 040Km 0 40km Source : INSEE, RP 2011, DGFIP, Impôt sur les revenus 2011. Calcul et cartographie : Céreq - ESO CNRS, Caen. LES ZONES À RISQUES D’ÉCHEC SCOLAIRE Diffi cultés moindres sur le plan économique et des diplômes Cumul de diffi cultés sur le plan familial, économique et des diplômes Diffi cultés prononcées sur le plan des diplômes et de l’emploi Diffi cultés sur le plan familial et des revenus 142 Mana Iracoubo Montsinéry-Tonnegrande Macouria Kourou St-Laurent- Cayenne du-Maroni Rémire-Montjoly Matoury Sinnamary Roura Approuage-Kaw Maripasoula Saint-Georges 040Km 0 40km Source : INSEE, RP 2011. Calcul et cartographie : Céreq - ESO CNRS, Caen. LA PART DES NON DIPLÔMÉS PARMI LES 15-24 ANS NON SCOLARISÉS 22,0 % à 31,4 % 31,4 % à 35,4 % 35,4 % à 41,1 % 41,1 % à 50,4 % 50,4 % à 63,0 % 63,0 % à 76,3 % 76,3 % à 92,1 % 143 ACADÉMIE DE GUYANE Le type « cumul de diffi cultés sur le plan familial, économique et des diplômes » ne représente que 4 cantons sur 14 mais concentre plus d’un habitant sur trois. La part des jeunes non diplômés parmi les 15-24 ans non scolarisés est parmi les plus élevées des DOM, hors Mayotte. Le type « diffi cultés sur le plan familial et des revenus » ne concerne que 2 cantons, mais ceux-ci concentrent encore plus les diffi cultés car ils regroupent 36 % de la population. La part des jeunes non diplômés parmi les 15-24 ans non scolarisés y est toutefois contenue dans la moyenne des DOM hors Mayotte. -

North Europe French Guiana North Brazil Exported on Saturday, October 02, 2021 at 22:51
North Europe French Guiana North Brazil Exported on Saturday, October 02, 2021 at 22:51 CMA CGM CMA CGM SAINT Vessel Name DOUCE FRANCE CMA CGM CAYENNE CMA CGM KOUROU CMA CGM MARSEILLE SINNAMARY LAURENT Voyage Ref. 0RT9ZS1MA 0RTA1S1MA 0RTA3S1MA 0RTA5S1MA 0RTA7S1MA 0RTA9S1MA Local Voyage Ref. ALGECIRAS Omitted Omitted Wednesday, September Omitted Omitted Wednesday, September 08, 2021 29, 2021 VIGO -- -- -- Thursday, September 16, Tuesday, September 21, -- 2021 2021 ROTTERDAM Thursday, September 02, -- Sunday, September 12, -- -- Sunday, October 03, 2021 2021 2021 LONDON GATEWAY Omitted Saturday, September 04, Tuesday, September 14, Monday, September 20, Saturday, September 25, Monday, October 04, 2021 PORT, ESS 2021 2021 2021 2021 LE HAVRE Sunday, September 05, Tuesday, September 07, Wednesday, September -- -- Tuesday, October 05, 2021 2021 2021 15, 2021 ROTTERDAM -- Omitted -- Tuesday, September 21, Sunday, September 26, -- 2021 2021 LE HAVRE -- -- -- Thursday, September 23, Tuesday, September 28, -- 2021 2021 PORT OF SPAIN Tuesday, September 14, -- -- -- -- -- 2021 PHILIPSBURG Omitted Friday, September 17, Friday, September 24, Saturday, October 02, 2021 Friday, October 08, 2021 Thursday, October 14, 2021 2021 2021 PORT OF SPAIN -- Sunday, September 19, Sunday, September 26, Sunday, October 03, 2021 Saturday, October 09, 2021 Saturday, October 16, 2021 2021 2021 Voyage Ref. 0RTA0N1MA 0RTA2N1MA 0RTA4N1MA 0RTA6N1MA 0RTA8N1MA 0RTAAN1MA Local Voyage Ref. DEGRAD DES CANNES Saturday, September 18, Wednesday, September Wednesday, September Thursday, -

From Lake Guaíba: Analysis of the Parasite Community
Parasitology Research https://doi.org/10.1007/s00436-018-5933-4 ORIGINAL PAPER Helminth fauna of Megaleporinus obtusidens (Characiformes: Anostomidae) from Lake Guaíba: analysis of the parasite community E. W. Wendt1 & C. M. Monteiro2 & S. B. Amato3 Received: 7 September 2017 /Accepted: 15 May 2018 # Springer-Verlag GmbH Germany, part of Springer Nature 2018 Abtract Structure of the helminth community of Megaleporinus obtusidens collected in Lake Guaíba was evaluated, and the results indicated that the diversity of helminth species was probably determined by fish behavior and eating habits. The influence of sex, weight, and standard length of hosts for parasitic indices was also analyzed. Sixteen helminth species were found parasitizing M. obtusidens, including the following: platyhelminths, with the highest richness, represented by one species of Aspidobothrea; four species of Digenea; and eight species of Monogenea; the latter, presented the highest prevalence. Rhinoxenus arietinus,foundin nasal cavities, had the greater abundance, and was the only species classified as core. The prevalence of Urocleidoides paradoxus was significantly influenced by the sex of the host; females had the highest values. Abundance was weakly influenced by fish weight and the body length of the hosts. Urocleidoides sp. had its abundance weakly influenced by the host weight. The other helminths were not influenced by biometric characteristics of the hosts. The total species richness was similar between male and female fish, and both had 14 helminth species of parasites. Keywords Host–parasite relationship . Fish biology . Lake environment . Southern Brazil Introduction It is found from north to south in Brazil, as well as in Argentina, Uruguay, and Paraguay (Britski et al. -

Synthèse Arrêté Préfectoral Du 25 Juin Portant Mesures De Prévention Et
Synthèse arrêté préfectoral du 25 juin portant mesures de prévention et restriction nécessaires dans le département de la Guyane dans le cadre de la lute contre le virus Covid 19. Article 1 : Maintien de l’interdiction de rassemblement de + de 10 personnes sauf motif professionnel , activité de voyage, ERP et cérémonie funéraire. Article 2 : Déplacements aux points de contrôle d’Iracoubo et Régina . Il faut justifier d’un motif familial ou personnel, santé ( si urgence), professionnel , approvisionnement en denrées. Le passage est soumis à contrôle médicalisé. ( je ne sais pas si cela signifie certificat médical. Je pense plutôt à une consultation médicale sur place comme à l’aeroport). Passage restreint : 8 h à 10 h et 13h à 15h en semaine . 8H à 11H samedi. Fermé dimanche. Cet article ne s’applique pas aux habitants de Iracoubo et Régina ( mais il faut un justificatif de domicile). Article 3 : Le déplacement des personnes à risque est strictement réglementé. La liste des personnes à risque est jointe. - Déplacements professionnels - Déplacements pour achats fournitures professionnelles ou de première nécessité . - Motifs de santé - Motif familial impérieux - Déplacement bref ( 1 heure et 1 km du domicile) - Convocation devant les tribunaux ou police /gendarmerie - Mission d’intérêt général sur demande de l’administration. CCI Guyane - Maison des Entreprises, Place de l’Esplanade | C.S 80049 | 97321 CAYENNE CEDEX T. 0594 29 96 00 | www.guyane.cci.fr Article 4 : Ces mêmes restrictions s’appliquent à toute la population de Saint Georges et de Camopi. Et pour sortir en dehors de la commune il faut l’un des 3 motifs impérieux. -

MOUVEMENT INTRA 2017 CERTIFIES / AGREGES Résultats Par Ordre Alphabétiques Des Collègues
MOUVEMENT INTRA 2017 CERTIFIES / AGREGES Résultats par ordre alphabétiques des collègues CODE TYPE BARE SPECIALIT NOM PRENOM ETABLISSEMENT MUTATION COMMUNE VŒUX ME E et L0080 ABUBAKARI SALIMAT CLG LA CANOPEE MATOURY 1GEO 77 L0080 BOCHER CAMILLE CLG JOSEPH HO TEN YOU KOUROU 2COM 27 L0080 EL JASEM HALIMA CLG EUGENIE TELL-EBOUE SAINT LAURENT DU MARONI 11ETA 46 B L7300 AGRICOLE ORPA NON MUTE L0100 LANDVELD CARLO LPO LEOPOLD ELFORT MANA 1ETAB 45 L0202 AMOUNIE GINA CLG PAUL JEAN-LOUIS SAINT LAURENT DU MARONI 1ETAB 102 L1500 BA MAMADOU ANNULATIION L0201 FUSEAU GERALDINE CLG AUGUSTE DEDE REMIRE-MONTJOLY 2COM 615,2 L0201 ILIC MIODRAG CLG AUXENCE CONTOUT CAYENNE 1ETAB 82 L0201 TRAVERS HELENE NON MUTE L0202 AINSEBA TAYEB CLG GERARD HOLDER CAYENNE 2ETAB 36 L0202 ALEXANDRE JEAN-ROLAND CLG ANTOINE SYLVERE FELIX MACOURIA 1ETAB 36 L1000 BERTIMON BEATRICE LGT LEON GONTRAN DAMAS REMIRE-MONTJOLY 1ETAB 82 L0202 ANDRIGHETTI SYLVIE CLG EUGENE NONNON CAYENNE 3ETAB 277 L0202 ANTHONI PATRICIA LPO LAMA PREVOT REMIRE-MONTJOLY 1ETAB 146 L0202 BLIN ANGELINA CLG LEO OTHILLY MANA 1ETAB 91 L0202 BROCQ BERNARD LPO LEOPOLD ELFORT MANA 1ETAB 111 L0202 BRUYERE FLORIAN CLG ANNEXE PIERRE ARDINET REGINA 3ETAB 27 L0202 CARBEL-FORTUNEE CAROLINE CLG GERARD HOLDER CAYENNE 3ETAB 27 L0202 CARLOTTI LUCILE CLG PAUL KAPEL CAYENNE 9GEO 27 L0202 CARMES NADIA CLG REEBERG NERON REMIRE-MONTJOLY 1COM 104 L0202 CHERY JOSILE NIXON LGT GASTON MONNERVILLE KOUROU 1ETAB 65 L0202 CISSE IBRAHIM LPO BERTENE JUMINER SAINT LAURENT DU MARONI 4GEO 65 L0202 COLIN PAMELA CLG PAUL JEAN-LOUIS SAINT LAURENT -

Alain Cornette La Ceramique Galibi En Guyane Française
ALAIN CORNETTE LA CERAMIQUE GALIBI EN GUYANE FRANÇAISE, ETUDE MORPHO-STYLISTIQUE ET TECHNIQUE RESUME Dans la région de la basse Mana, on trouve encore quelques potières dans certains villages galibi ("Carib côtiers", Amérindiens de la famille linguistique karib). Bien que supplantée par l'aluminium ou les matières plastiques, la poterie traditionnelle survit, assurant une fonction de plus en plus décorative au détriment de l'utilitaire. Cette étude utilise les données provenant de sources archéologiques (fouilles et analyses), ethnographiques (nomenclature galibi, récits et légendes) et bibliographiques, d'une part; ethnographiques (observation directe de chaque étape de la fabrication, et analyse), d'autre part. Le recoupement de ces données, exploitées statistiquement, devra permettre la détermination des caractéristiques de cette céramique. Ce corpus de données servira ainsi de référence, à la fois technique et chronologique, en vue d'une étude générale de la céramique amérindienne provenant de certains secteurs archéologiques de la Guyane, en particulier sur le littoral. OBJECTIFS DE CE TRAVAIL Cette étude comporte deux parties: - la première concerna la céramique galibi1 elle-même; - la seconde s'intéresse à une comparaison avec d'autres céramiques amérindien nes de la Guyane, en utilisant les données de la première. Il s'agit donc, dans un premier temps, d'essayer de bien cerner la céramique des Indiens Galibi vivant actuellement en Guyane, en prenant en compte simultanément les aspects morphologiques et morphométriques, stylistiques, techniques et fonctionnels. Ce travail se base sur l'étude des objets qui sera complétée par une observation directe de leur fabrication. En exploitant statistiquement cet ensemble de données, nous mènerons une étude comparative avec les céramiques provenant des sites archéologiques de différents secteurs, notamment la basse Mana, la bande littorale entre Saint-Laurent- du-Maroni et Cayenne, et Nle-de-Cayenne.