An Update of the Revision of Cyathea Subgen. Alsophila Sect
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Dfggfdgfdgsdfsdfdsfdsfsdfsdfdsfs
dfggfdgfdgsdfsdfdsfdsfsdfsdfdsfsdfdsfdmmm REPOBLIKAN'I MADAGASIKARA Fitiavana - Tanindrazana - Fandrosoana ----------------- HAUTE COUR CONSTITUTIONNELLE RESULTATS DEFINITIFS DU SECOND TOUR DE L'ELECTION PRESIDENTIELLE DU 19 DECEMBRE 2018 dfggfdgffhCode BV: 330601010101 dfggfdgffhBureau de vote: EPP ALAKAMISY ITENINA SALLE 1 dfggfdgffhCommune: ALAKAMISY ITENINA dfggfdgffhDistrict: VOHIBATO dfggfdgffhRegion: HAUTE MATSIATRA dfggfdgffhProvince: FIANARANTSOA Inscrits : 405 Votants: 238 Blancs et Nuls: 2 Soit: 0,84% Suffrages exprimes: 236 Soit: 99,16% Taux de participation: 58,77% N° d'ordre Logo Photo Nom et Prenoms Candidat Voix obtenues Pourcentage 13 RAJOELINA Andry Nirina 92 38,98% 25 RAVALOMANANA Marc 144 61,02% Total voix: 236 100,00% Copyright @ HCC 2019 dfggfdgfdgsdfsdfdsfdsfsdfsdfdsfsdfdsfdmmm REPOBLIKAN'I MADAGASIKARA Fitiavana - Tanindrazana - Fandrosoana ----------------- HAUTE COUR CONSTITUTIONNELLE RESULTATS DEFINITIFS DU SECOND TOUR DE L'ELECTION PRESIDENTIELLE DU 19 DECEMBRE 2018 dfggfdgffhCode BV: 330601020101 dfggfdgffhBureau de vote: EPP AMBALAFAHIBATO SALLE 1 dfggfdgffhCommune: ALAKAMISY ITENINA dfggfdgffhDistrict: VOHIBATO dfggfdgffhRegion: HAUTE MATSIATRA dfggfdgffhProvince: FIANARANTSOA Inscrits : 576 Votants: 212 Blancs et Nuls: 3 Soit: 1,42% Suffrages exprimes: 209 Soit: 98,58% Taux de participation: 36,81% N° d'ordre Logo Photo Nom et Prenoms Candidat Voix obtenues Pourcentage 13 RAJOELINA Andry Nirina 68 32,54% 25 RAVALOMANANA Marc 141 67,46% Total voix: 209 100,00% Copyright @ HCC 2019 dfggfdgfdgsdfsdfdsfdsfsdfsdfdsfsdfdsfdmmm -

Small Hydro Resource Mapping in Madagascar
Public Disclosure Authorized Small Hydro Resource Mapping in Madagascar INCEPTION REPORT [ENGLISH VERSION] August 2014 Public Disclosure Authorized Public Disclosure Authorized Public Disclosure Authorized This report was prepared by SHER Ingénieurs-Conseils s.a. in association with Mhylab, under contract to The World Bank. It is one of several outputs from the small hydro Renewable Energy Resource Mapping and Geospatial Planning [Project ID: P145350]. This activity is funded and supported by the Energy Sector Management Assistance Program (ESMAP), a multi-donor trust fund administered by The World Bank, under a global initiative on Renewable Energy Resource Mapping. Further details on the initiative can be obtained from the ESMAP website. This document is an interim output from the above-mentioned project. Users are strongly advised to exercise caution when utilizing the information and data contained, as this has not been subject to full peer review. The final, validated, peer reviewed output from this project will be a Madagascar Small Hydro Atlas, which will be published once the project is completed. Copyright © 2014 International Bank for Reconstruction and Development / THE WORLD BANK Washington DC 20433 Telephone: +1-202-473-1000 Internet: www.worldbank.org This work is a product of the consultants listed, and not of World Bank staff. The findings, interpretations, and conclusions expressed in this work do not necessarily reflect the views of The World Bank, its Board of Executive Directors, or the governments they represent. The World Bank does not guarantee the accuracy of the data included in this work and accept no responsibility for any consequence of their use. -

1 COAG No. 72068718CA00001
COAG No. 72068718CA00001 1 TABLE OF CONTENT I- EXECUTIVE SUMMARY .................................................................................................................................................. 6 II- INTRODUCTION ....................................................................................................................................................... 10 III- MAIN ACHIEVEMENTS DURING QUARTER 1 ........................................................................................................... 10 III.1. IR 1: Enhanced coordination among the public, nonprofit, and commercial sectors for reliable supply and distribution of quality health products ........................................................................................................................... 10 III.2. IR2: Strengthened capacity of the GOM to sustainably provide quality health products to the Malagasy people 15 III.3. IR 3: Expanded engagement of the commercial health sector to serve new health product markets, according to health needs and consumer demand ........................................................................................................ 36 III.4. IR 4: Improved sustainability of social marketing to deliver affordable, accessible health products to the Malagasy people ............................................................................................................................................................. 48 III.5. IR5: Increased demand for and use of health products among the Malagasy people -

Ma D a G a S C
M A D A G A S C A R A W o r l d A p a r t S a t 0 8 N o v e m b e r - T u e 2 5 N o v e m b e r 2 0 1 4 Formerly a landlocked plateau in the centre of Gondwanaland, Madagascar became marooned when that ancient landmass split into the island continents of Australia, Antarctica, South America and Africa. Separated by the Mozambique Channel from Africa’s eastern coast about 65 million years ago, Madagascar developed in isolation. Its natural history is unique. The world’s fourth‐largest island is another Galápagos, referred to by some ecologists as ‘the eighth continent’. Eighty percent of Malagasy plants and animals are endemic, rivalling Brazil in its biodiversity. Archaeologists believe that people began to arrive in Madagascar from Indonesia/Malaya about 4000 years ago and although they have eliminated some species they haven’t dominated nature: there’s simply too much of it. Eight whole plant families exist only on Madagascar, as do over 1000 orchid species, many thousands of succulents, countless insects, at least 350 species of frog, around 400 kinds of reptile, five families of birds and approaching 215 different non‐marine mammals including lemurs, the prosimians who comprise an entire branch of the primate family tree, the order to which we ourselves belong. Australian Museum Members offer interesting and worthwhile travel experiences. Madagascar will never be a low cost mass tourism destination: but it will be one you will never forget. -

Sustainable Landscapes in Eastern Madagascar Environmental And
Sustainable Landscapes in Eastern Madagascar Environmental and Social Management Plan Translation of the original French version 19 May 2016 (Updated 23 August 2016) 1 Table of Contents Acronyms ............................................................................................................................................... 7 Glossary ................................................................................................................................................. 9 Executive Summary ............................................................................................................................ 10 1 Introduction ................................................................................................................................. 17 1.1 Background and Project Objectives ...................................................................................... 17 1.2 Objectives of the ESMP ........................................................................................................ 17 1.3 Link between the ESMP and the Environmental and Social Management Tools for the COFAV and CAZ Protected Areas ........................................................................................................ 18 2 Project Overview ......................................................................................................................... 20 2.1 Description of Components, Activities, and Relevant Sectors .............................................. 20 2.2 Targets and Characteristics -

Madagascar Guided Walking Adventure
Natural Wonders of Madagascar Tour Style: Worldwide Multi-Centre Destination: Madagascar Trip code: MCWHW Trip Walking Grade: 2 HOLIDAY OVERVIEW Immerse yourself in the culture and unique biodiversity of this incredible island. Trek through the impressive canyons and sheer sandstone cliffs that jut upwards across Isalo National Park and marvel at the cascading waterfalls of Andringitra National Park. Wander through gorgeous landscapes and the lush, dense rainforests of Ranomafana National Park. Nature lovers can discover the unique wildlife of one of the world’s most diverse ecosystems, including endearing lemur species, colourful chameleons and beautiful orchid blooms. WHAT'S INCLUDED • 12 nights’ accommodation in en-suite rooms, 2 nights in a tented camp • Full board, from breakfast on day 2 to dinner on day 15 • Full holiday programme with a specialist local guide • Internal flight, national park entrance fees and travel by air-conditioned minibus or coach • Tips www.hfholidays.co.uk PAGE 1 [email protected] Tel: +44(0) 20 3974 8865 HOLIDAYS HIGHLIGHTS • Explore the national parks of this enigmatic island. Visit Isalo, Andringitra and Ranomafana • Snap shots of Madagascar’s many endemic species • Enjoy walks through rainforests, traditional villages and magnificent mountains • Discover magical Malagasy culture • Option to extend the tour to visit Andasibe National Park TRIP SUITABILITY This holiday is at the upper end of our level 2 walks. Trails are rough and rocky underfoot. Walks are 3-6½ miles (5-10km) with up to 1,850 feet (570m) of ascent. There is quite a lot of travelling on this tour, on some poorly made roads. -

Commune Sahambavy – District Lalangina Region Haute Matsiatra
ETUDES DE CAS -COMMUNE SAHAMBAVY – DISTRICT LALANGINA REGION HAUTE MATSIATRA « Le genre et la lutte contre la violence comme levier de développement » Informations sur la commune PAYS MADAGASCAR COMMUNE SAHAMBAVY Score de base 67 Dernier score 67 Nombre de population 18 522 dont 9 413 femmes et 9 109 hommes Caractéristiques de la commune La cueillette des feuilles de thé Personne contact RANDRIANIRINA Aimé Robert Fonction Adjoint au Maire Coordonnées Téléphone: 032 28 266 28 E-mail : [email protected] La commune rurale de Sahambavy se trouve à 22 kilomètres à l'est de Fianarantsoa. Elle est également traversée par le chemin de fer reliant Fianarantsoa à Manakara. Sahambavy se démarque surtout par son champ théicole - unique à Madagascar - ainsi que son lac artificiel devenu un lieu d'attraction. En 2011, il n’y avait qu’une seule femme parmis le personnel de la commune, le genre a été exclue du programme. On a constaté la faible participation des femmes dans le processus de développement, elles n’osent pas participer aux différentes réunions publiques et aux formations. Les femmes n’arrivent pas à denoncer les actes de violences. Depuis la signature de la convention entre la commune et le programme UADEL/ACORDS, la commune de Sahambavy a commencé à faire des sensibilisations et vulgarisations de l’approche genre au niveau de cette commune, différents événements ont été organisés notamment les renforcements des capacités et mise en place des différentes strucutures. Le programme SAHA a pris le relais dans les étapes suivantes: la prise en compte des femmes vulnérables, la valorisation des problématiques féminines, l’implication des jeunes dans le processus de développement local. -
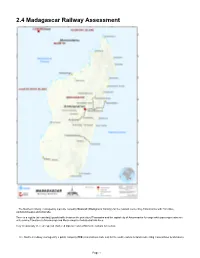
2.4 Madagascar Railway Assessment
2.4 Madagascar Railway Assessment - The Northern railway, managed by a private company Madarail (Madagascar Railway) for the network connecting Antananarivo with Tamatave, Ambatondrazaka and Antsirabe. There is a regular (at least daily) goods traffic between the port city of Toamasina and the capital city of Antananarivo for cargo while passenger trains are only serving Tamatave to Moramanga and Moramanga to Ambatrodrazaka lines. Very occasionally there are special chartered trips on restored Micheline railcars for tourists. - The Southern railway, managed by a public company FCE (Fianarantsoa Cote Est) for the south eastern network connecting Fianarantsoa to Manakara. Page 1 The southern line has regular passenger and cargo trains, which provides a slow but picturesque alternative to the recently rehabilitated road in the region. For more information on railway company contact details, please see the following link: Madagascar Railway Assessment Railway Companies and Consortia 4.2.7 Madagascar Railway Company Contact List Northern railway*: *During our study, Madarail was in the midst of restructuring, therefore, they did not want to share information, statistics or even contacts. All the information gathered and shared in this document comes exclusively from third parties or from data found on the internet. Madarail, was founded on October 10, 2002 following the decision of the Malagasy State to privatize the Malagasy National Railway Network1 (RNCFM). A concession agreement for the management of the North network is then established between the new private operator and the State. Madarail began operating the Northern railway network in Madagascar on 1 July 2003. In 2008, the Belgian operator Vecturis, already active in eight other African countries, became the majority shareholder of the company and the new railway operator. -

Universite D'antananarivo ******** ********
UNIVERSITE D’ANTANANARIVO ******** FACULTE DE DROIT, D’ECONOMIE, DE GESTION ET DE SOCIOLOGIE ******** FORMATION PROFESIONNALISANTE EN TRAVAIL SOCIAL ET DEVELOPPEMENT MEMOIRE DE FIN D’ETUDE EN VUE DE L’OBTENTION DU DIPLOME DE LICENCE PROFESSIONNELLE EN TRAVAIL SOCIAL ET DEVELOPPEMENT DIAGNOSTIC SOCIO-ECONOMIQUE DES EXPLOITATIONS PISCICOLES CAS DE L’ASSOCIATION APPAFI COMMUNE RURALE DE TALATA AMPANO (REGION HAUTE MATSIATRA) Présenté par : Mlle MANATSAFIDY Sambatra Rovatina Membres du jury : Président : Monsieur ETIENNE Stefano Raherimalala, Maitre de conférences Juge : Monsieur SAMUEL Richard, AESR Encadreur pédagogique : Pr SOLOFOMIARANA RAPANOEL Bruno Allain Encadreur professionnel : Monsieur RASOJA Paul Année universitaire : 2015-2016 Soutenu le lundi 21 Mars 2016 à 11h DIAGNOSTIC SOCIO-ECONOMIQUE DES EXPLOITATIONS PISCICOLES CAS DE L’ASSOCIATION APPAFI COMMUNE DE TALATA AMPANO (REGION HAUTE MATSIATRA) i SOMMAIRE INTRODUCTION GENERALE Ier PARTIE : PRESENTATION DE LA ZONE D’ETUDE ET ANCRAGE THEORIQUE CHAPITRE 1. ETAT DES LIEUX CHAPITRE 2. REPERES THEORICO-CONCEPTUEL ET METHODOLOGIQUE IIème PARTIE : PRESENTATION DES RESULTATS ET OPERATIONNALISATION DES HYPOTHESES CHAPITRE 3. INVENTAIRE SOCIOLOGIQUE DE LA POPULATION CIBLE CHAPITRE 4. EVALUATION DE L’ACTIVITE PISCICOLE DE LA POPULATION CIBLE PARTIE III : DISCUSSIONS ET APPROCHE PROSPECTIVES CHAPITRE 5. ANALYSE DES PROBLEMES AUTOUR DE LA FILIERE ET VERIFICATION DES HYPOTHESES CHAPITRE 6. DISCUSSIONS ET REFLEXIONS PROSPECTIVES CONCLUSION GENERALE BIBLIOGRAPHIE Table des matières Liste des -

USAID/Madagascar and Community Health Volunteers: Working in Partnership to Achieve Health Goals
USAID/Madagascar and Community Health Volunteers: Working in Partnership to Achieve Health Goals USAID/MADAGASCAR AND COMMUNITY HEALTH VOLUNTEERS: WORKING IN PARTNERSHIP TO ACHIEVE HEALTH GOALS CONTENTS INTRODUCTION .............................................................................................................................................1 Supporting the Health Sector and CHVs in Madagascar ........................................................... 2 CHV Package of Services .................................................................................................................. 3 Impact of CHVs on Health Care .....................................................................................................4 SUCCESS STORIES: INNOVATIONS, COMMUNITY ENGAGEMENT, AND SUSTAINABILITY ................................................................................................................................... 5 Innovations ........................................................................................................................................... 5 Community Engagement ................................................................................................................... 7 Sustainability .........................................................................................................................................9 CHALLENGES ..................................................................................................................................................13 LESSONS -

Rapport D'activites 2003
Association Humanitaire Akamasoa BP : 7010 – Antananarivo – 101 – Madagascar RAPPORT D’ACTIVITES 2003 1. Rappel des objectifs, des lieux d’intervention et des personnes soutenues 1.1. Objectif principal L’association humanitaire « Akamasoa » a été créée par le Père Pedro OPEKA en 1989. Elle a pour but de contribuer à la réhabilitation humaine et la réinsertion économique et sociale de 3.420 familles (15.560 personnes dont 8.400 enfants scolarisés) issues des rues d’Antananarivo, des banlieues de cette capitale et de familles de la Province de Fianarantsoa. Cette action humanitaire consiste à accueillir de manière permanente ces familles dans les centres créés par l’association. Cet accueil permanent est indispensable pour assurer la longue éducation des adultes pour la citoyenneté et l’instruction des enfants et des adolescents. 1.2. Objectifs spécifiques a) Faire accéder les adultes à un travail rémunéré, soit par l’association soit à l’extérieur, pour qu’ils subviennent à leurs charges, en particulier le logement familial et l’accès aux services de santé créés par l’association. b) Assurer l’éducation scolaire obligatoire des enfants et des adolescents de la classe primaire au baccalauréat dans les écoles et lycées créés par l’association ; établissements reconnus par l’Etat. c) Consolider l’accueil d’urgence temporaire de familles démunies vivant dans les rues et sans travail par la fourniture d’aides en nature : soins sanitaires, aide alimentaire de secours et dons d’outils de travail de base. 1.3. Lieux d’intervention Province d’Antananarivo 1. Centre d’Antolojanahary (créé en 1989) - RN 4, PK 60 - route d’Ankazobe 2. -

2. Literature Review
2. LITERATURE REVIEW Geology The geology of the RNP region, and most of eastern Madagascar, consists of Precambrian metamorphic and igneous basement rocks (Figures 2.1 and 2.2). This Precambrian basement testifies to a complex geologic history that researchers have tentatively deciphered only within the last decade. The recent geologic advances have implications for pedology research in eastern Madagascar, because despite the availability of relatively high-quality geologic maps from the 1950s-1970s, no well- supported structural/tectonic scenario has previously existed for systematically correlating the igneous-metamorphic sequences from one region to another (Windley et al., 1994; Kroner et al., 2000). These geologic correlations provide a basis for more reliably defining the “soil systems” (Buol et al., 1989; Daniels et al., 1999) of the RNP region and possibly other areas of the eastern escarpment zone. Until recently, even the origin of Madagascar's current geographic position was a source of lively scientific debate. Boast and Nairn (1982) summarized the three then- prevailing theories as to Madagascar's pre-Jurassic paleoposition: 1) adjacent to the Somalia-Kenya-Tanzania coast, 2) next to the Mozambique-Natal coast, or 3) essentially located where the island occurs today. A broad consensus has since emerged that Madagascar's paleoposition was adjacent to the present-day Somalia- Kenya-Tanzania coast, based upon a variety of data sources including magnetic anomalies, paleomagnetic data, geochronology, structural trends, lithologic correlations, stratigraphy, bathymetry, and biogeography (Martelat et al. et al., 1997, and sources therein; Paquette and Nédeléc, 1998; PALEOMAP Project, 1999). 11 RNP* Figure 2.1. Simplified geology map by Du Puy and Moat (1996), after Besairie (1964).