Selective Oxidation of Carbolide C–H Bonds by an Engineered Macrolide P450 Mono-Oxygenase
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

First Principles Prediction of Thermodynamic Properties
2 First Principles Prediction of Thermodynamic Properties Hélio F. Dos Santos and Wagner B. De Almeida NEQC: Núcleo de Estudos em Química Computacional, Departamento de Química, ICE Universidade Federal de Juiz de Fora (UFJF), Campus Universitário Martelos, Juiz de Fora LQC-MM: Laboratório de Química Computacional e Modelagem Molecular Departamento de Química, ICEx, Universidade Federal de Minas Gerais (UFMG) Campus Universitário, Pampulha, Belo Horizonte Brazil 1. Introduction The determination of the molecular structure is undoubtedly an important issue in chemistry. The knowledge of the tridimensional structure allows the understanding and prediction of the chemical-physics properties and the potential applications of the resulting material. Nevertheless, even for a pure substance, the structure and measured properties reflect the behavior of many distinct geometries (conformers) averaged by the Boltzmann distribution. In general, for flexible molecules, several conformers can be found and the analysis of the physical and chemical properties of these isomers is known as conformational analysis (Eliel, 1965). In most of the cases, the conformational processes are associated with small rotational barriers around single bonds, and this fact often leads to mixtures, in which many conformations may exist in equilibrium (Franklin & Feltkamp, 1965). Therefore, the determination of temperature-dependent conformational population is very much welcomed in conformational analysis studies carried out by both experimentalists and theoreticians. -

New Chemistry of Donor-Acceptor Cycloalkanes and Studies Towards the Synthesis of Streptorubin B
Western University Scholarship@Western Electronic Thesis and Dissertation Repository 8-2-2016 12:00 AM New Chemistry of Donor-Acceptor Cycloalkanes and Studies Towards the Synthesis of Streptorubin B Naresh Vemula The University of Western Ontario Supervisor Prof. Brian L. Pagenkopf The University of Western Ontario Graduate Program in Chemistry A thesis submitted in partial fulfillment of the equirr ements for the degree in Doctor of Philosophy © Naresh Vemula 2016 Follow this and additional works at: https://ir.lib.uwo.ca/etd Part of the Organic Chemistry Commons Recommended Citation Vemula, Naresh, "New Chemistry of Donor-Acceptor Cycloalkanes and Studies Towards the Synthesis of Streptorubin B" (2016). Electronic Thesis and Dissertation Repository. 3895. https://ir.lib.uwo.ca/etd/3895 This Dissertation/Thesis is brought to you for free and open access by Scholarship@Western. It has been accepted for inclusion in Electronic Thesis and Dissertation Repository by an authorized administrator of Scholarship@Western. For more information, please contact [email protected]. Abstract and Keywords Abstract This dissertation presents two separate chapters within the broad area of synthetic organic chemistry. The first chapter describes the annelation chemistry of donor-acceptor (DA) cyclopropanes and cyclobutanes for the synthesis of heterocycles. The Yb(OTf)3-catalyzed [4+2] cycloaddition between DA cyclobutanes and nitrosoarenes facilitated the synthesis of tetrahydro-1,2-oxazines in good to excellent yields as single diastereomers. Additionally, an unexpected deoxygenation occurred with electron-rich nitrosoarenes under MgI2-catalysis that afforded pyrrolidine products. The GaCl3-catalyzed [4+2] cycloaddition of DA cyclobutanes and cis-diazenes provided hexahydropyridazine derivatives in good to excellent yields as single diastereomers. -

Toxicological Assessment of Hanford Tank Headspace Chemicals - Determination of Chemicals of Potential Concern
PNNL-14949 Rev. 0 Pacific Northwest National Laboratory Operated by Battelle for the U.S. Department of Energy Toxicological Assessment of Hanford Tank Headspace Chemicals - Determination of Chemicals of Potential Concern I. E. Burgeson N. A. Moore A. L. Bunn J. L. Huckaby December 2004 • Prepared for CH2M HILL Hanford Group, Inc. LEGAL NOTICE This report was prepared by Battelle Memorial Institute (Battelle) as an account of sponsored research activities. Neither Client nor Battelle nor any person acting on behalf of either: MAKES ANY WARRANTY OR REPRESENTATION, EXPRESS OR IMPLIED, with respect to the accuracy, completeness, or usefulness of the information contained in this report, or that the use of any information, apparatus, process, or composition disclosed in this report may not infringe privately owned rights; or Assumes any liabilities with respect to the use of, or for damages resulting from the use of, any information, apparatus, process, or composition disclosed in this report. Reference herein to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise, does not necessarily constitute or imply its endorsement, recommendation, or favoring by Battelle. The views and opinions of authors expressed herein do not necessarily state or reflect those of Battelle. @ This document was printed on recycled paper. (9/97) PNNL-14949 Rev. 0 Toxicological Assessment of Hanford Tank Headspace Chemicals - Determination of Chemicals of Potential Concern I. E. Burgeson N. A. Moore A. L. Bunn J. L. Huckaby December 2004 Prepared for CH2M HILL Hanford Group, Inc. Pacific Northwest Nationa] Laboratory Richland, Washington 99352 Summary A toxicological assessment of chemicals found, or predicted to be present, in Hanford tank headspaces was performed by Pacific Northwest National Laboratory in support of the Industrial Hygiene Chemical Vapor Technical Basis produced by CH2M HILL Hanford Group, Inc. -

Cycloalkanes, Cycloalkenes, and Cycloalkynes
CYCLOALKANES, CYCLOALKENES, AND CYCLOALKYNES any important hydrocarbons, known as cycloalkanes, contain rings of carbon atoms linked together by single bonds. The simple cycloalkanes of formula (CH,), make up a particularly important homologous series in which the chemical properties change in a much more dramatic way with increasing n than do those of the acyclic hydrocarbons CH,(CH,),,-,H. The cyclo- alkanes with small rings (n = 3-6) are of special interest in exhibiting chemical properties intermediate between those of alkanes and alkenes. In this chapter we will show how this behavior can be explained in terms of angle strain and steric hindrance, concepts that have been introduced previously and will be used with increasing frequency as we proceed further. We also discuss the conformations of cycloalkanes, especially cyclo- hexane, in detail because of their importance to the chemistry of many kinds of naturally occurring organic compounds. Some attention also will be paid to polycyclic compounds, substances with more than one ring, and to cyclo- alkenes and cycloalkynes. 12-1 NOMENCLATURE AND PHYSICAL PROPERTIES OF CYCLOALKANES The IUPAC system for naming cycloalkanes and cycloalkenes was presented in some detail in Sections 3-2 and 3-3, and you may wish to review that ma- terial before proceeding further. Additional procedures are required for naming 446 12 Cycloalkanes, Cycloalkenes, and Cycloalkynes Table 12-1 Physical Properties of Alkanes and Cycloalkanes Density, Compounds Bp, "C Mp, "C diO,g ml-' propane cyclopropane butane cyclobutane pentane cyclopentane hexane cyclohexane heptane cycloheptane octane cyclooctane nonane cyclononane "At -40". bUnder pressure. polycyclic compounds, which have rings with common carbons, and these will be discussed later in this chapter. -
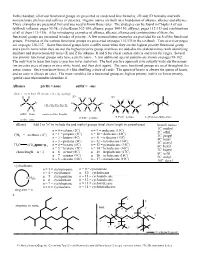
In This Handout, All of Our Functional Groups Are Presented As Condensed Line Formulas, 2D and 3D Formulas and with Nomenclature Prefixes and Suffixes (If Present)
In this handout, all of our functional groups are presented as condensed line formulas, 2D and 3D formulas and with nomenclature prefixes and suffixes (if present). Organic names are built on a foundation of alkanes, alkenes and alkynes. Those examples are presented first and you need to know those rules. The strategies can be found in Chapter 4 of our textbook (alkanes: pages 93-98, cycloalkanes 102-104, alkenes: pages 104-110, alkynes: pages 112-113 and combinations of all of them 113-115). After introducing examples of alkanes, alkenes, alkynes and combinations of them, the functional groups are presented in order of priority. A few nomenclature examples are provided for each of the functional groups. Examples of the various functional groups are presented on pages 115-135 in the textbook. Two overview pages are on pages 136-137. Some functional groups have a suffix name when they are the highest priority functional group and a prefix name when they are not the highest priority group, and these are added to the skeletal names with identifying numbers and stereochemistry terms (E and Z for alkenes, R and S for chiral centers and cis and trans for rings). Several low priority functional groups only have a prefix name. A few additional special patterns are shown on pages 98-102. The only way to learn this topic is practice (over and over). The best practice approach is to actually write out the names (on an extra piece of paper or on a white board, and then do it again). The same functional groups are used throughout the entire course. -

Conversion of Waste Plastic Into Liquid Hydrocarbons (ENERGY) by Cuco 3 Catalyst: Application of Scientific Research on Plastic Pollution
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by International Institute for Science, Technology and Education (IISTE): E-Journals Chemical and Process Engineering Research www.iiste.org ISSN 2224-7467 (Paper) ISSN 2225-0913 (Online) Vol.48, 2017 Conversion of Waste Plastic into Liquid Hydrocarbons (ENERGY) by Cuco 3 Catalyst: Application of Scientific Research on Plastic Pollution Manvir Singh 1 Sudesh Kumar 1 Moinuddin Sarker 2 1.Department of Chemistry Banasthali University Rajasthan india 2.Waste Technologies LLC, Department of Research & Development, 1376 Chopsey Hill Road, Bridgeport, CT- 06606, USA Abstract Waste plastics were converted into valuable liquid hydrocarbon fuel. it is can be used as different purpose of energy-source such as petrol engines, diesel engines, generators, vehicles and its good source of chemicals etc. Plastics have many properties like light weight, high durability so its demand increases in every sector. Pyrolysis of the waste plastic (hdpe) was carried out with CuCO 3 catalysts and temperature range from 0 °C to 390 °C. The collected liquid hydrocarbons fuel was characterized by FT-IR, NMR, GCxGCMS spectrometer and fuel density was 78 g ml -1 and conversion was very good. The research paper exhibits concentrating on application for solving daily life issues and problems of plastic. Keywords: Pyrolysis, Liquid hydrocarbons fuel, CuCO 3 catalysts, Conversion, Glass reactor, GCxGCMS. Introduction First time plastic was invented by Alexander Parkes in 1862 that has a high molecular weight (Brydson 1999). A molecule formed by repetition of simple units is called polymer. Plastic is also called polymer for example polyethylene (Chanda 2000). -

Cyclohexane: a Strain-Free Cycloalkane
Organic Chemistry I Mohammad Jafarzadeh Faculty of Chemistry, Razi University Organic Chemistry, Structure and Function (7th edition) By P. Vollhardt and N. Schore, Elsevier, 2014 1 CYCLOHEXANE: A STRAIN-FREE CYCLOALKANE The chair conformation of cyclohexane is strain free A hypothetical planar cyclohexane would suffer from 12 H–H eclipsing interactions and sixfold bond-angle strain (a regular hexagon requires 120o bond angles). One conformation of cyclohexane, obtained by moving carbons 1 and 4 out of planarity in opposite directions, is in fact strain free. This structure is called the chair conformation of cyclohexane (because it resembles a chair), in which eclipsing is completely absent, and the bond angles are very nearly tetrahedral. o -1 The calculated �H comb of cyclohexane (-944.4 kcal mol ) based on a strain-free (CH2)6 model is very close to the experimentally determined value (-944.5 kcal mol-1). Indeed, the C–C bond strength, �Ho = 88 kcal mol-1 (368 kJ mol-1), is normal. The molecular model of cyclohexane enables to recognize the conformational stability of the molecule. If we view it along (any) one C–C bond, we can see the staggered arrangement of all substituent groups along it. Because of its lack of strain, cyclohexane is as inert as a straight-chain alkane. 2 140 CHAPTER 4 Cycloalkanes 140 CHAPTER 4 Cycloalkanes Model Building 4-3 CYCLOHEXANE: A STRAIN-FREE CYCLOALKANE Model Building The cyclohexane4-3 ringCYCLOHEXANE: is one of the most common A STRAIN-FREE and important CYCLOALKANE structural units in organic chemistry. Its substituted derivatives exist in many natural products (see Section 4-7), and an understandingThe cyclohexane of its conformational ring is one of mobility the most is ancommon important and aspect important of organic structural chemistry. -

Support Based Cobalt Catalysts for Liquid Hydrocarbon Fuel Production from Syngas
Supplementary Alumina Coated Silica Nanosprings (NS) Support Based Cobalt Catalysts for Liquid Hydrocarbon Fuel Production From Syngas Abdulbaset Alayat, Elena Echeverria, Farid Sotoudehniakarani, David N. Mcllroy and Armando G. McDonald * Table S1. The FT products identified by GC-MS for unmodified Co/NS catalyst reduced by H2 at a temperature of 230 °C and H2/CO = 2. Retention Time (min) Identified Compounds Mol(%) M+ (m/z) 2.16 2-pentene 3-methyl 0.65 84 2.21 3-hexene-(z) 0.72 84 2.29 2-ethyl-1-pentanol 2.09 116 2.38 3-Methylhexane 0.45 100 2.56 cis-1,2 -dimethyl cyclopentane 7.4 98 2.65 Heptane 1.83 100 2.75 3-Heptene 1.21 98 2.77 Isopropylcyclobutane 1.0 98 2.88 Cycloheptane 0.68 98 3.35 Cyclopropane, (1,2-dimethylpropyl) 0.13 112 3.41 3-Methyl-1-heptene 0.78 112 3.54 5-Methyl-1-heptene 1.65 112 3.69 6-Methyl-1-heptene 1.38 130 3.71 1-octanol 1.24 130 3.84 3-Ethyl-4-methyl-1-pentanol 1.24 130 4.12 3,4-Dimethyl-1-hexene 0.38 112 4.20 Cyclopropane, pentyl 5.72 112 4.38 Octane 2.05 114 4.56 1,2-Diethylcyclobutane 0.76 112 4.75 2-Octene 0.48 112 5.54 Cis-3-Nonene 0.52 126 5.75 2,3-Diethyl-3-heptene 1.68 126 5.95 6-Methyl-1-octene 1.34 126 6.16 4-Ethylheptane 1.17 128 6.39 2-Nonene, (E)- 0.27 126 6.57 7-Methyl-1-octene 0.41 126 6.70 1-Nonene 3.37 126 6.93 Nonane 1.62 128 7.13 2-Nonene, (E)- 0.75 126 7.38 Cis-4-Nonene 0.68 126 7.90 2,3,5-Trimethylhexane 0.26 128 8.33 Cyclopropane, 1-methyl-2-pentyl- 0.49 140 8.48 1-Decene 1.09 140 8.74 3,7-Dimethyl-1-octanol 1.09 158 9.01 3-Methylnonane 0.48 142 9.21 cis-4-Decene 0.46 140 9.48 (Z)-2-Decene -

Chromene Compound Chromenverbindung Composé Chromène
(19) TZZ_ ¥_T (11) EP 1 657 239 B1 (12) EUROPEAN PATENT SPECIFICATION (45) Date of publication and mention (51) Int Cl.: of the grant of the patent: C07D 311/94 (2006.01) C07D 405/10 (2006.01) 11.08.2010 Bulletin 2010/32 C07D 409/04 (2006.01) C07D 451/02 (2006.01) C07D 453/02 (2006.01) C07D 491/20 (2006.01) (2006.01) (2006.01) (21) Application number: 06000884.4 C07D 491/107 C07D 495/10 C09K 9/02 (2006.01) G03C 1/73 (2006.01) G02B 1/04 (2006.01) C07D 491/10 (2006.01) (22) Date of filing: 28.12.2000 C07D 493/10 (2006.01) (54) Chromene compound Chromenverbindung Composé chromène (84) Designated Contracting States: • Takeda, Yasuko DE ES FR GB IT Shunan-shi Yamaguchi 745-0053 (JP) (30) Priority: 21.02.2000 JP 2000042682 • Momoda, Junji Shunan-shi (43) Date of publication of application: Yamaguchi 745-0053 (JP) 17.05.2006 Bulletin 2006/20 • Nagoh, Hironobu Shunan-shi (62) Document number(s) of the earlier application(s) in Yamaguchi 745-0053 (JP) accordance with Art. 76 EPC: 00985992.7 / 1 184 379 (74) Representative: Benson, John Everett et al J. A. Kemp & Co. (73) Proprietor: Tokuyama Corporation 14 South Square Shunan-shi, Yamaguchi-ken 745-8648 (JP) Gray’s Inn London WC1R 5JJ (GB) (72) Inventors: • Izumi, Shinobu (56) References cited: Shunan-shi EP-A- 1 054 010 WO-A-00/71544 Yamaguchi 745-0053 (JP) WO-A-01/02384 DE-A- 19 902 771 • Kawabata, Yuichiro JP-A- 2000 344 762 US-A- 6 113 814 Shunan-shi Yamaguchi 745-0053 (JP) Note: Within nine months of the publication of the mention of the grant of the European patent in the European Patent Bulletin, any person may give notice to the European Patent Office of opposition to that patent, in accordance with the Implementing Regulations. -

Introduction (Pdf)
Dictionary of Natural Products on CD-ROM This introduction screen gives access to (a) a general introduction to the scope and content of DNP on CD-ROM, followed by (b) an extensive review of the different types of natural product and the way in which they are organised and categorised in DNP. You may access the section of your choice by clicking on the appropriate line below, or you may scroll through the text forwards or backwards from any point. Introduction to the DNP database page 3 Data presentation and organisation 3 Derivatives and variants 3 Chemical names and synonyms 4 CAS Registry Numbers 6 Diagrams 7 Stereochemical conventions 7 Molecular formula and molecular weight 8 Source 9 Importance/use 9 Type of Compound 9 Physical Data 9 Hazard and toxicity information 10 Bibliographic References 11 Journal abbreviations 12 Entry under review 12 Description of Natural Product Structures 13 Aliphatic natural products 15 Semiochemicals 15 Lipids 22 Polyketides 29 Carbohydrates 35 Oxygen heterocycles 44 Simple aromatic natural products 45 Benzofuranoids 48 Benzopyranoids 49 1 Flavonoids page 51 Tannins 60 Lignans 64 Polycyclic aromatic natural products 68 Terpenoids 72 Monoterpenoids 73 Sesquiterpenoids 77 Diterpenoids 101 Sesterterpenoids 118 Triterpenoids 121 Tetraterpenoids 131 Miscellaneous terpenoids 133 Meroterpenoids 133 Steroids 135 The sterols 140 Aminoacids and peptides 148 Aminoacids 148 Peptides 150 β-Lactams 151 Glycopeptides 153 Alkaloids 154 Alkaloids derived from ornithine 154 Alkaloids derived from lysine 156 Alkaloids -

Using JCP Format
Estimating Solid–Liquid Phase Change Enthalpies and Entropies Cite as: Journal of Physical and Chemical Reference Data 28, 1535 (1999); https:// doi.org/10.1063/1.556045 Submitted: 07 January 1999 . Published Online: 28 April 2000 James S. Chickos, William E. Acree, and Joel F. Liebman ARTICLES YOU MAY BE INTERESTED IN Phase Transition Enthalpy Measurements of Organic and Organometallic Compounds. Sublimation, Vaporization and Fusion Enthalpies From 1880 to 2010 Journal of Physical and Chemical Reference Data 39, 043101 (2010); https:// doi.org/10.1063/1.3309507 Heat Capacities and Entropies of Organic Compounds in the Condensed Phase. Volume III Journal of Physical and Chemical Reference Data 25, 1 (1996); https://doi.org/10.1063/1.555985 Enthalpies of Vaporization of Organic and Organometallic Compounds, 1880–2002 Journal of Physical and Chemical Reference Data 32, 519 (2003); https:// doi.org/10.1063/1.1529214 Journal of Physical and Chemical Reference Data 28, 1535 (1999); https://doi.org/10.1063/1.556045 28, 1535 © 1999 American Institute of Physics and American Chemical Society. Estimating Solid–Liquid Phase Change Enthalpies and Entropies James S. Chickosa… Department of Chemistry, University of Missouri–St. Louis, St. Louis Missouri 63121 William E. Acree, Jr. Department of Chemistry, University of North Texas, Denton, Texas 76203 Joel F. Liebman Department of Chemistry and Biochemistry, University of Maryland Baltimore County, Baltimore, Maryland 21250 Received January 7, 1999; revised manuscript received July 10, 1999 A group additivity method based on molecular structure is described that can be used ⌬Tfus ⌬Tfus to estimate solid–liquid total phase change entropy ( 0 Stpce) and enthalpy ( 0 Htpce) of organic molecules. -

Epa Region 2
ENVIRONMENTAL PROTECTION AGENCY REGION 5 ELECTRONIC DATA DELIVERABLE VALID VALUES REFERENCE MANUAL Appendix to EPA Electronic Data Deliverable (EDD) Comprehensive Specification Manual . Aug, 2016 ELECTRONIC DATA DELIVERABLE VALID VALUES REFERENCE MANUAL Appendix to EPA Electronic Data Deliverable (EDD) Comprehensive Specification Manual TABLE OF CONTENTS Table A-1 Matrix .......................................................................................................................................... 5 Table A-2 Coord Geometric type ................................................................................................................. 7 Table A-3 Horizontal Collection Method ..................................................................................................... 7 Table A-4 Horizontal Accuracy Units .......................................................................................................... 8 Table A-5 Horizontal Datum ........................................................................................................................ 8 Table A-6 Elevation Collection Method ....................................................................................................... 8 Table A-7 Elevation Datum .......................................................................................................................... 9 Table A-8 Material ........................................................................................................................................ 9 Table A-9