Middle Triassic Sharks from the Catalan Coastal Ranges (NE Spain) and Faunal Colonization Patterns During the Westward Transgression of Tethys T ⁎ E
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

JVP 26(3) September 2006—ABSTRACTS
Neoceti Symposium, Saturday 8:45 acid-prepared osteolepiforms Medoevia and Gogonasus has offered strong support for BODY SIZE AND CRYPTIC TROPHIC SEPARATION OF GENERALIZED Jarvik’s interpretation, but Eusthenopteron itself has not been reexamined in detail. PIERCE-FEEDING CETACEANS: THE ROLE OF FEEDING DIVERSITY DUR- Uncertainty has persisted about the relationship between the large endoskeletal “fenestra ING THE RISE OF THE NEOCETI endochoanalis” and the apparently much smaller choana, and about the occlusion of upper ADAM, Peter, Univ. of California, Los Angeles, Los Angeles, CA; JETT, Kristin, Univ. of and lower jaw fangs relative to the choana. California, Davis, Davis, CA; OLSON, Joshua, Univ. of California, Los Angeles, Los A CT scan investigation of a large skull of Eusthenopteron, carried out in collaboration Angeles, CA with University of Texas and Parc de Miguasha, offers an opportunity to image and digital- Marine mammals with homodont dentition and relatively little specialization of the feeding ly “dissect” a complete three-dimensional snout region. We find that a choana is indeed apparatus are often categorized as generalist eaters of squid and fish. However, analyses of present, somewhat narrower but otherwise similar to that described by Jarvik. It does not many modern ecosystems reveal the importance of body size in determining trophic parti- receive the anterior coronoid fang, which bites mesial to the edge of the dermopalatine and tioning and diversity among predators. We established relationships between body sizes of is received by a pit in that bone. The fenestra endochoanalis is partly floored by the vomer extant cetaceans and their prey in order to infer prey size and potential trophic separation of and the dermopalatine, restricting the choana to the lateral part of the fenestra. -

Post-Carboniferous Stratigraphy, Northeastern Alaska by R
Post-Carboniferous Stratigraphy, Northeastern Alaska By R. L. DETTERMAN, H. N. REISER, W. P. BROSGE,and]. T. DUTRO,JR. GEOLOGICAL SURVEY PROFESSIONAL PAPER 886 Sedirnentary rocks of Permian to Quaternary age are named, described, and correlated with standard stratigraphic sequences UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON 1975 UNITED STATES DEPARTMENT OF THE INTERIOR ROGERS C. B. MORTON, Secretary GEOLOGICAL SURVEY V. E. McKelvey, Director Library of Congress Cataloging in Publication Data Detterman, Robert L. Post-Carboniferous stratigraphy, northeastern Alaska. (Geological Survey Professional Paper 886) Bibliography: p. 45-46. Supt. of Docs. No.: I 19.16:886 1. Geology-Alaska. I. Detterman, Robert L. II. Series: United States. Geological Survey. Professional Paper 886. QE84.N74P67 551.7'6'09798 74-28084 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 Stock Number 024-001-02687-2 CONTENTS Page Page Abstract __ _ _ _ _ __ __ _ _ _ _ _ _ _ _ _ _ _ _ __ __ _ _ _ _ _ _ __ __ _ _ __ __ __ _ _ _ _ __ 1 Stratigraphy__:_Continued Introduction __________ ----------____ ----------------____ __ 1 Kingak Shale ---------------------------------------- 18 Purpose and scope ----------------------~------------- 1 Ignek Formation (abandoned) -------------------------- 20 Geographic setting ------------------------------------ 1 Okpikruak Formation (geographically restricted) ________ 21 Previous work and acknowledgments ------------------ 1 Kongakut Formation ---------------------------------- -

OFR21 a Guide to Fossil Sharks, Skates, and Rays from The
STATE OF DELAWARE UNIVERSITY OF DELAWARE DELAWARE GEOLOGICAL SURVEY OPEN FILE REPORT No. 21 A GUIDE TO FOSSIL SHARKS J SKATES J AND RAYS FROM THE CHESAPEAKE ANU DELAWARE CANAL AREA) DELAWARE BY EDWARD M. LAUGINIGER AND EUGENE F. HARTSTEIN NEWARK) DELAWARE MAY 1983 Reprinted 6-95 FOREWORD The authors of this paper are serious avocational students of paleontology. We are pleased to present their work on vertebrate fossils found in Delaware, a subject that has not before been adequately investigated. Edward M. Lauginiger of Wilmington, Delaware teaches biology at Academy Park High School in Sharon Hill, Pennsyl vania. He is especially interested in fossils from the Cretaceous. Eugene F. Hartstein, also of Wilmington, is a chemical engineer with a particular interest in echinoderm and vertebrate fossils. Their combined efforts on this study total 13 years. They have pursued the subject in New Jersey, Maryland, and Texas as well as in Delaware. Both authors are members of the Mid-America Paleontology Society, the Delaware Valley Paleontology Society, and the Delaware Mineralogical Society. We believe that Messrs. Lauginiger and Hartstein have made a significant technical contribution that will be of interest to both professional and amateur paleontologists. Robert R. Jordan State Geologist A GUIDE TO FOSSIL SHARKS, SKATES, AND RAYS FROM THE CHESAPEAKE AND DELAWARE CANAL AREA, DELAWARE Edward M. Lauginiger and Eugene F. Hartstein INTRODUCTION In recent years there has been a renewed interest by both amateur and professional paleontologists in the rich upper Cretaceous exposures along the Chesapeake and Delaware Canal, Delaware (Fig. 1). Large quantities of fossil material, mostly clams, oysters, and snails have been collected as a result of this activity. -

USGS Geologic Investigations Series I-2673, Pamphlet
U.S. DEPARTMENT OF THE INTERIOR TO ACCOMPANY MAP I–2673 U.S. GEOLOGICAL SURVEY GEOLOGIC MAP OF THE ARCTIC QUADRANGLE, ALASKA By W.P. Brosgé, H.N. Reiser, J.T. Dutro, Jr., R.L. Detterman, and I.L. Tailleur INTRODUCTION parts of the quadrangle in 1926 and 1927 and published his results in 1930 (Mertie, 1930). This rapid reconnaissance, GEOLOGIC SETTING AND LAND USE made mostly on foot under extremely difficult conditions, sketched out the rudimentary distribution of rock types, but The Arctic quadrangle is well located to shed light on little of the modern stratigraphy was discerned. The distri- the basic geologic relations of northern Alaska. The rocks bution of the Lisburne Limestone in the eastern part of the represent all of the stratigraphic systems from Cambrian to quadrangle was roughly shown; and the approximate distri- Cretaceous and all but one of the tectono-stratigraphic bution of the Devonian clastic rocks between the East Fork subterranes of the Brooks Range, from the autochthonous of the Chandalar and the Sheenjek valley was also shown subterrane in the north to the allochthonous subterranes far- although details of the history of these strata were not known. ther south. Most of the pre-Mississippian carbonate rocks were mapped Among the distinctive geologic features displayed in the by Mertie as Skajit Limestone, supposedly of Silurian ages. Arctic quadrangle are voluminous volcanic rocks of prob- He mapped the chert and slate unit between Old Woman able Devonian age, a wide array of Carboniferous carbonate Creek and the East Fork of the Chandalar, and tentatively facies in the Lisburne Group (which here extends up into the assigned it a Mississippian or Late Devonian age. -

The Rhaetian Vertebrates of Chipping Sodbury, South Gloucestershire, UK, a Comparative Study
Lakin, R. J., Duffin, C. J., Hildebrandt, C., & Benton, M. J. (2016). The Rhaetian vertebrates of Chipping Sodbury, South Gloucestershire, UK, a comparative study. Proceedings of the Geologists' Association, 127(1), 40-52. https://doi.org/10.1016/j.pgeola.2016.02.010 Peer reviewed version License (if available): Unspecified Link to published version (if available): 10.1016/j.pgeola.2016.02.010 Link to publication record in Explore Bristol Research PDF-document This is the author accepted manuscript (AAM). The final published version (version of record) is available online via Elsevier at http://www.sciencedirect.com/science/article/pii/S0016787816000183. Please refer to any applicable terms of use of the publisher. University of Bristol - Explore Bristol Research General rights This document is made available in accordance with publisher policies. Please cite only the published version using the reference above. Full terms of use are available: http://www.bristol.ac.uk/red/research-policy/pure/user-guides/ebr-terms/ *Manuscript Click here to view linked References 1 The Rhaetian vertebrates of Chipping Sodbury, South Gloucestershire, UK, 1 2 3 a comparative study 4 5 6 7 8 Rebecca J. Lakina, Christopher J. Duffinaa,b,c, Claudia Hildebrandta, Michael J. Bentona 9 10 a 11 School of Earth Sciences, University of Bristol, BS8 1RJ, UK 12 13 b146 Church Hill Road, Sutton, Surrey, SM3 8NF, UK. 14 15 c 16 Earth Sciences Department, The Natural History Museum, Cromwell Road, London, SW7 17 18 5BD, UK. 19 20 21 22 23 ABSTRACT 24 25 Microvertebrates are common in the basal bone bed of the Westbury Formation of England, 26 27 28 documenting a fauna dominated by fishes that existed at the time of the Rhaetian 29 30 Transgression, some 206 Myr ago. -

Revision Des Faunes De Vertebres Du Site De Proven Cheres-Sur-Meuse (Trias Terminal, Nord-Est De La France)
REVISION DES FAUNES DE VERTEBRES DU SITE DE PROVEN CHERES-SUR-MEUSE (TRIAS TERMINAL, NORD-EST DE LA FRANCE) par Gilles CUNY * SOMMAIRE Page Résumé, Abstract ..................... : . .. 103 Introduction ..................................................................... 103 Historique. 103 Révision des anciennes collections ................................................... 105 Muséum National d'Histoire Naturelle ................... 105 Université Pierre et Marie Curie. .. .. .. 106 Besançon . 120 Dijon. .. .. .. .. 121 Langres - Saint-Dizier. 121 Lyon ....................................................................... 123 Etude du matériel récent. 126 Discussion ...................................................................... 127 Conclusion ................................................................. :. 129 Remerciements. .. 130 Bibliographie . .. 130 Légendes des planches. 134 * Laboratoire de Paléontologie des Vertébrés, Université Pierre et Marie Curie - Boîte 106, 4 place Jussieu, 75252 PARIS cédex OS, France. Mots-clés: Trias, Rhétien, Poissons, Amphibiens, Reptiles Key-words: Triassic, Rhetian, Fishes, Amphibians, Reptiles PalaeoveT/ebra/a. Montpellier, 24 (1-2): 10H34. 6 fig .• 3 pl. (Reçu le 23 Février 1994. accepté le 15 Mai 1994. publié le 14 Juin 1995) -~----------------- - --.------- --------------- 500MHRES -- ------ - ------ -- --- ---- -------- ---.- A--- ..L -L ~ ~ 1.10 1.00 1 <6~::<:<=.: :~:~:::,'EE: J '_ 0.50 =...- ~ ::_-_____ -___ ---___ ~~:: ~-- __ .- _______ ~:: ----___ ~: ~-____ : ~ ~-. ~ ~ '_____ -

The Early Triassic Jurong Fish Fauna, South China Age, Anatomy, Taphonomy, and Global Correlation
Global and Planetary Change 180 (2019) 33–50 Contents lists available at ScienceDirect Global and Planetary Change journal homepage: www.elsevier.com/locate/gloplacha Research article The Early Triassic Jurong fish fauna, South China: Age, anatomy, T taphonomy, and global correlation ⁎ Xincheng Qiua, Yaling Xua, Zhong-Qiang Chena, , Michael J. Bentonb, Wen Wenc, Yuangeng Huanga, Siqi Wua a State Key Laboratory of Biogeology and Environmental Geology, China University of Geosciences (Wuhan), Wuhan 430074, China b School of Earth Sciences, University of Bristol, BS8 1QU, UK c Chengdu Center of China Geological Survey, Chengdu 610081, China ARTICLE INFO ABSTRACT Keywords: As the higher trophic guilds in marine food chains, top predators such as larger fishes and reptiles are important Lower Triassic indicators that a marine ecosystem has recovered following a crisis. Early Triassic marine fishes and reptiles Fish nodule therefore are key proxies in reconstructing the ecosystem recovery process after the end-Permian mass extinc- Redox condition tion. In South China, the Early Triassic Jurong fish fauna is the earliest marine vertebrate assemblage inthe Ecosystem recovery period. It is constrained as mid-late Smithian in age based on both conodont biostratigraphy and carbon Taphonomy isotopic correlations. The Jurong fishes are all preserved in calcareous nodules embedded in black shaleofthe Lower Triassic Lower Qinglong Formation, and the fauna comprises at least three genera of Paraseminotidae and Perleididae. The phosphatic fish bodies often show exceptionally preserved interior structures, including net- work structures of possible organ walls and cartilages. Microanalysis reveals the well-preserved micro-structures (i.e. collagen layers) of teleost scales and fish fins. -

Copyrighted Material
06_250317 part1-3.qxd 12/13/05 7:32 PM Page 15 Phylum Chordata Chordates are placed in the superphylum Deuterostomia. The possible rela- tionships of the chordates and deuterostomes to other metazoans are dis- cussed in Halanych (2004). He restricts the taxon of deuterostomes to the chordates and their proposed immediate sister group, a taxon comprising the hemichordates, echinoderms, and the wormlike Xenoturbella. The phylum Chordata has been used by most recent workers to encompass members of the subphyla Urochordata (tunicates or sea-squirts), Cephalochordata (lancelets), and Craniata (fishes, amphibians, reptiles, birds, and mammals). The Cephalochordata and Craniata form a mono- phyletic group (e.g., Cameron et al., 2000; Halanych, 2004). Much disagree- ment exists concerning the interrelationships and classification of the Chordata, and the inclusion of the urochordates as sister to the cephalochor- dates and craniates is not as broadly held as the sister-group relationship of cephalochordates and craniates (Halanych, 2004). Many excitingCOPYRIGHTED fossil finds in recent years MATERIAL reveal what the first fishes may have looked like, and these finds push the fossil record of fishes back into the early Cambrian, far further back than previously known. There is still much difference of opinion on the phylogenetic position of these new Cambrian species, and many new discoveries and changes in early fish systematics may be expected over the next decade. As noted by Halanych (2004), D.-G. (D.) Shu and collaborators have discovered fossil ascidians (e.g., Cheungkongella), cephalochordate-like yunnanozoans (Haikouella and Yunnanozoon), and jaw- less craniates (Myllokunmingia, and its junior synonym Haikouichthys) over the 15 06_250317 part1-3.qxd 12/13/05 7:32 PM Page 16 16 Fishes of the World last few years that push the origins of these three major taxa at least into the Lower Cambrian (approximately 530–540 million years ago). -
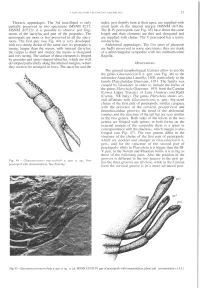
Thoracic Appendages. the 3Rd Maxilliped Is Only Partially
A NEW DECAPOD CRUSTACEAN ASSEMBLAGE 11 Thoracic appendages. The 3rd maxilliped is only index, just slightly bent at their apex, are supplied with partially preserved in two specimens (MSNB 8237; small teeth on the internal margin (MSNM i 10736). MSNM i 10732): it is possible to observe just frag- The II-IV pereiopods (see Fig. 45) have about the same ments of the dactylus and part of the propodus. The length and their elements are thin and elongated and pereiopods are more or less preserved in all the speci- are supplied with chelae. The V pereiopod has a termi- mens. The first pair (see Fig. 44) is very developed, nal dactylus. with two strong chelae of the same size; its propodus is Abdominal appendages. The five pairs of pleopods strong, longer than the merus, with internal dactylus; are badly preserved in some specimens; they are made the carpus is short and stocky; the merus is elongated of a subrectangular sympodite with two multiarticulate and very strong. The surface of these elements is fringed flagella. by granules and spine-shaped tubercles, which are well developed particularly along the internal margins, where Observations they seem to be arranged in rows. The dactylus and the The general morphological features allow to ascribe the genus Glaessnericaris n. gen. (see Fig. 46) to the infraorder Astacidea Latreille, 1803, particularly to the family Platychelidae Glaessner, 1931. The family was created by Glaessner in order to include the forms of the genus Platychela Glaessner, 1931 from the Carnian (Lower Upper Triassic) of Lunz (Austria) and Raibl (Carnia, NE Italy). -

The Rocks in the Iberian Basin …………………………….………………
The Permian-Triassic rocks of the eastern Iberian Ranges: A general approach in the context of the first stages of the break-up of Pangea. Pan- European Correlation of the Triassic. 10th International Field Workshop. 1 2 2 1,2 Raúl De la Horra , José López-Gómez , Alfredo Arche , María José Escudero-Mozo , Belén Galán-Abellán1,2, José F. Barrenechea2,3, Javier Martín-Chivelet1,2, and Violeta Borruel2 With collaborations of: Ana Márquez-Aliaga4, Pablo Plasencia5, Cristina Pla5, Leopoldo Márquez5, 6 Marceliano Lago (1)Departamento de Estratigrafía. Facultad de Ciencias Geológicas. Universidad Complutense de Madrid. 28040 Madrid, Spain. [email protected]; [email protected]; [email protected]; [email protected] (2)Instituto de Geociencias IGEO (CSIC,UCM), c/ José Antonio Nováis 2, 28040 Madrid, Spain. [email protected]; [email protected] (3)Departamento de Cristalografía y Mineralogía. Facultad de Ciencias Geológicas. Universidad Complutense de Madrid. 28040 Madrid, Spain. [email protected] (4) Instituto Cavanilles de Biodiversidad y Biología Evolutiva. Departamento de Geología de la Universidad de Valencia, 46100 Burjassot, Valencia, Spain. [email protected] (5) Departamento de Geología de la Universidad de Valencia, 46100 Burjassot, Valencia, Spain. (6) Departamento de Ciencias de la Tierra, Universidad de Zaragoza, 50.009 Zaragoza, Spain. [email protected] Reservados todos los derechos. Ni la totalidad ni parte de este libro puede reproducirse, almacenarse o transmitirse en materia alguna por ningún medio sin permiso por escrito del Instituto de Geociencias IGEO (CSIC-UCM). The Permian-Triassic rocks of the eastern Iberian Ranges: A general approach in the context of the first stages of the break-up of Pangea. -

Database of Bibliography of Living/Fossil
www.shark-references.com Version 16.01.2018 Bibliography database of living/fossil sharks, rays and chimaeras (Chondrichthyes: Elasmobranchii, Holocephali) Papers of the year 2017 published by Jürgen Pollerspöck, Benediktinerring 34, 94569 Stephansposching, Germany and Nicolas Straube, Munich, Germany ISSN: 2195-6499 DOI: 10.13140/RG.2.2.32409.72801 copyright by the authors 1 please inform us about missing papers: [email protected] www.shark-references.com Version 16.01.2018 Abstract: This paper contains a collection of 817 citations (no conference abstracts) on topics related to extant and extinct Chondrichthyes (sharks, rays, and chimaeras) as well as a list of Chondrichthyan species and hosted parasites newly described in 2017. The list is the result of regular queries in numerous journals, books and online publications. It provides a complete list of publication citations as well as a database report containing rearranged subsets of the list sorted by the keyword statistics, extant and extinct genera and species descriptions from the years 2000 to 2017, list of descriptions of extinct and extant species from 2017, parasitology, reproduction, distribution, diet, conservation, and taxonomy. The paper is intended to be consulted for information. In addition, we provide data information on the geographic and depth distribution of newly described species, i.e. the type specimens from the years 1990 to 2017 in a hot spot analysis. New in this year's POTY is the subheader "biodiversity" comprising a complete list of all valid chimaeriform, selachian and batoid species, as well as a list of the top 20 most researched chondrichthyan species. Please note that the content of this paper has been compiled to the best of our abilities based on current knowledge and practice, however, possible errors cannot entirely be excluded. -

Morphometric Discriminant Analysis of Isolated Chondrichthyan Scales for Palaeoecological Inferences: the Middle Triassic Of
Journal of Iberian Geology 40 (1) 2014: 87-97 http://dx.doi.org/10.5209/rev_JIGE.2014.v40.n1.44089 www.ucm.es /info/estratig/journal.htm ISSN (print): 1698-6180. ISSN (online): 1886-7995 Morphometric Discriminant Analysis of isolated chondrichthyan scales for palaeoecological inferences: the Middle Triassic of the Iberian Chain (Spain) as a case of study H. Ferrón1, C. Pla1, C. Martínez-Pérez1,2, M.J. Escudero-Mozo3,4, H. Botella1* 1Departamento de Geología, Universidad de Valencia, Avda. Dr. Moliner, 50, 46100 Burjassot, Spain. 2School of Earth Sciences, University of Bristol, Wills Memorial Building, Queen’s Road, Bristol BS8 1RJ, United Kingdom. 3 Departamento de Estratigrafía, Universidad Complutense de Madrid, C/José Antonio Novais 12, 28040 Madrid, Spain 4 Instituto de Geociencias IGEO (CSIC,UCM), C/José Antonio Novais 12, 28040 Madrid, Spain e-mail addresses: [email protected] (H.F.), [email protected] (C.P.), [email protected] (C.M.-P.); [email protected] (M.J.E.-M.) [email protected] (H.B., *corresponding author) Received: 12 December 2012 / Accepted: 4 December 2013 / Available online: 25 February 2014 Abstract Palaeontological studies on exosqueletal disarticulated remains of chondrichthyans have focused on teeth and only less interest has been paid to scales due their limited taxonomic and systematic significance. However, classical works linking the morphology and the function of the squamation in extant sharks suggest that, despite their limited taxonomic value, the study of isolated scales can be a useful tool for palaeoenvironmental and palaeoecological inferences. Following this idea, we have analyzed the fossil record of shark scales from two Middle Triassic sections of the Iberian Chain (Spain), identifying different functional types by means of a morphometric discriminant analysis.