Morphometric Discriminant Analysis of Isolated Chondrichthyan Scales for Palaeoecological Inferences: the Middle Triassic Of
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Revision Des Faunes De Vertebres Du Site De Proven Cheres-Sur-Meuse (Trias Terminal, Nord-Est De La France)
REVISION DES FAUNES DE VERTEBRES DU SITE DE PROVEN CHERES-SUR-MEUSE (TRIAS TERMINAL, NORD-EST DE LA FRANCE) par Gilles CUNY * SOMMAIRE Page Résumé, Abstract ..................... : . .. 103 Introduction ..................................................................... 103 Historique. 103 Révision des anciennes collections ................................................... 105 Muséum National d'Histoire Naturelle ................... 105 Université Pierre et Marie Curie. .. .. .. 106 Besançon . 120 Dijon. .. .. .. .. 121 Langres - Saint-Dizier. 121 Lyon ....................................................................... 123 Etude du matériel récent. 126 Discussion ...................................................................... 127 Conclusion ................................................................. :. 129 Remerciements. .. 130 Bibliographie . .. 130 Légendes des planches. 134 * Laboratoire de Paléontologie des Vertébrés, Université Pierre et Marie Curie - Boîte 106, 4 place Jussieu, 75252 PARIS cédex OS, France. Mots-clés: Trias, Rhétien, Poissons, Amphibiens, Reptiles Key-words: Triassic, Rhetian, Fishes, Amphibians, Reptiles PalaeoveT/ebra/a. Montpellier, 24 (1-2): 10H34. 6 fig .• 3 pl. (Reçu le 23 Février 1994. accepté le 15 Mai 1994. publié le 14 Juin 1995) -~----------------- - --.------- --------------- 500MHRES -- ------ - ------ -- --- ---- -------- ---.- A--- ..L -L ~ ~ 1.10 1.00 1 <6~::<:<=.: :~:~:::,'EE: J '_ 0.50 =...- ~ ::_-_____ -___ ---___ ~~:: ~-- __ .- _______ ~:: ----___ ~: ~-____ : ~ ~-. ~ ~ '_____ -

Copyrighted Material
06_250317 part1-3.qxd 12/13/05 7:32 PM Page 15 Phylum Chordata Chordates are placed in the superphylum Deuterostomia. The possible rela- tionships of the chordates and deuterostomes to other metazoans are dis- cussed in Halanych (2004). He restricts the taxon of deuterostomes to the chordates and their proposed immediate sister group, a taxon comprising the hemichordates, echinoderms, and the wormlike Xenoturbella. The phylum Chordata has been used by most recent workers to encompass members of the subphyla Urochordata (tunicates or sea-squirts), Cephalochordata (lancelets), and Craniata (fishes, amphibians, reptiles, birds, and mammals). The Cephalochordata and Craniata form a mono- phyletic group (e.g., Cameron et al., 2000; Halanych, 2004). Much disagree- ment exists concerning the interrelationships and classification of the Chordata, and the inclusion of the urochordates as sister to the cephalochor- dates and craniates is not as broadly held as the sister-group relationship of cephalochordates and craniates (Halanych, 2004). Many excitingCOPYRIGHTED fossil finds in recent years MATERIAL reveal what the first fishes may have looked like, and these finds push the fossil record of fishes back into the early Cambrian, far further back than previously known. There is still much difference of opinion on the phylogenetic position of these new Cambrian species, and many new discoveries and changes in early fish systematics may be expected over the next decade. As noted by Halanych (2004), D.-G. (D.) Shu and collaborators have discovered fossil ascidians (e.g., Cheungkongella), cephalochordate-like yunnanozoans (Haikouella and Yunnanozoon), and jaw- less craniates (Myllokunmingia, and its junior synonym Haikouichthys) over the 15 06_250317 part1-3.qxd 12/13/05 7:32 PM Page 16 16 Fishes of the World last few years that push the origins of these three major taxa at least into the Lower Cambrian (approximately 530–540 million years ago). -
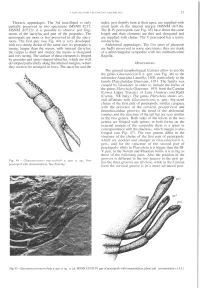
Thoracic Appendages. the 3Rd Maxilliped Is Only Partially
A NEW DECAPOD CRUSTACEAN ASSEMBLAGE 11 Thoracic appendages. The 3rd maxilliped is only index, just slightly bent at their apex, are supplied with partially preserved in two specimens (MSNB 8237; small teeth on the internal margin (MSNM i 10736). MSNM i 10732): it is possible to observe just frag- The II-IV pereiopods (see Fig. 45) have about the same ments of the dactylus and part of the propodus. The length and their elements are thin and elongated and pereiopods are more or less preserved in all the speci- are supplied with chelae. The V pereiopod has a termi- mens. The first pair (see Fig. 44) is very developed, nal dactylus. with two strong chelae of the same size; its propodus is Abdominal appendages. The five pairs of pleopods strong, longer than the merus, with internal dactylus; are badly preserved in some specimens; they are made the carpus is short and stocky; the merus is elongated of a subrectangular sympodite with two multiarticulate and very strong. The surface of these elements is fringed flagella. by granules and spine-shaped tubercles, which are well developed particularly along the internal margins, where Observations they seem to be arranged in rows. The dactylus and the The general morphological features allow to ascribe the genus Glaessnericaris n. gen. (see Fig. 46) to the infraorder Astacidea Latreille, 1803, particularly to the family Platychelidae Glaessner, 1931. The family was created by Glaessner in order to include the forms of the genus Platychela Glaessner, 1931 from the Carnian (Lower Upper Triassic) of Lunz (Austria) and Raibl (Carnia, NE Italy). -

Shark) Dental Morphology During the Early Mesozoic Dynamik Av Selachian (Haj) Tandmorfologi Under Tidig Mesozoisk
Examensarbete vid Institutionen för geovetenskaper Degree Project at the Department of Earth Sciences ISSN 1650-6553 Nr 399 Dynamics of Selachian (Shark) Dental Morphology During the Early Mesozoic Dynamik av Selachian (haj) tandmorfologi under Tidig Mesozoisk Alexander Paxinos INSTITUTIONEN FÖR GEOVETENSKAPER DEPARTMENT OF EARTH SCIENCES Examensarbete vid Institutionen för geovetenskaper Degree Project at the Department of Earth Sciences ISSN 1650-6553 Nr 399 Dynamics of Selachian (Shark) Dental Morphology During the Early Mesozoic Dynamik av Selachian (haj) tandmorfologi under Tidig Mesozoisk Alexander Paxinos ISSN 1650-6553 Copyright © Alexander Paxinos Published at Department of Earth Sciences, Uppsala University (www.geo.uu.se), Uppsala, 2017 Abstract Dynamics of Selachian (Shark) Dental Morphology During the Early Mesozoic Alexander Paxinos The ancestors of all modern day sharks and rays (Neoselachii) may have appeared during the Late Palaeozoic, but their major diversification happened sometime during the Early Mesozoic. Taxonomic evidence places the first neoselachian diversification in the Early Jurassic. Taxonomic diversity analyses, however, are often affected by incompleteness of the fossil record and sampling biases. On the other hand, the range of morphological variation (disparity) offers a different perspective for studying evolutionary patterns across time. Disparity analyses are much more resilient to sampling biases than diversity analyses, and disparity has the potential to provide a more ecologically-relevant context. -

Middle Triassic Sharks from the Catalan Coastal Ranges (NE Spain) and Faunal Colonization Patterns During the Westward Transgression of Tethys T ⁎ E
Palaeogeography, Palaeoclimatology, Palaeoecology 539 (2020) 109489 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Middle Triassic sharks from the Catalan Coastal ranges (NE Spain) and faunal colonization patterns during the westward transgression of Tethys T ⁎ E. Manzanaresa, M.J. Escudero-Mozob, H. Ferrónc,d, C. Martínez-Pérezc,d, H. Botellac, a Botany and Geology Department, University of Valencia, Avda. Dr. Moliner, 50, 46100 Burjassot, Valencia, Spain b Instituto de Geociencias, UCM, CSIC, Calle del Dr. Severo Ochoa, 7, 28040 Madrid, Spain c Institut Cavanilles de Biodiversitat i Biologia Evolutiva, Universitat de Valencia, Paterna 46980, Valencia, Spain d School of Earth Sciences, University of Bristol, Bristol BS8 1TQ, UK ARTICLE INFO ABSTRACT Keywords: Palaeogeographic changes that occurred during the Middle Triassic in the westernmost Tethyan domain were Dispersal strategies governed by a westward marine transgression of the Tethys Ocean. The transgression flooded wide areas of the Palaeocurrents eastern part of Iberia, forming new epicontinental shallow-marine environments, which were subsequently Anisian colonized by diverse faunas, including chondrichthyans. The transgression is recorded by two successive Ladinian transgressive–regressive cycles: (1) middle–late Anisian and (2) late Anisian–early Carnian. Here, we describe Coastal chondrichthyans the chondrichthyan fauna recovered from several Middle Triassic stratigraphic sections (Pelsonian- Longobardian) located at the Catalan Coastal Basin (western-most Tethys). The assemblage consists of isolated teeth of the species Hybodus plicatilis, Omanoselache bucheri, O. contrarius and Pseudodalatias henarejensis. Our data complement a series of recent studies on chondrichthyan faunas from Middle-Late Triassic marine basins of the Iberian Peninsula, allowing us to evaluate patterns of faunal colonization. -

Database of Bibliography of Living/Fossil
www.shark-references.com Version 09.01.2012 Bibliography database of living/fossil sharks, rays and chimaeras (Chondrichtyes: Elasmobranchii, Holocephali) Papers of the year 2009 published by Jürgen Pollerspöck, Benediktinerring 34, 94569 Stephansposching, Germany ISSN: 2195-6499 - 1 - please inform me about missing papers E-Mail: [email protected] www.shark-references.com Version 09.01.2012 Abstract: This collection is the result of research in numerous journals, books and online publications. It contains more than 500 citations of papers about living/fossil sharks, rays and chimaeras (Chondrichtyes: Elasmobranchii, Holocephali). Notice: This paper is intended to be consulted for advice and information. This information has been compiled to the best of my abilities based on current knowledge and practice, however, please note that possible errors cannot be altogether/entirely excluded. Citation: Pollerspöck, J. (2011), Bibliography database of living/fossil sharks, rays and chimaeras (Chondrichtyes: Elasmobranchii, Holocephali) - Papers of the year 2009 -, www.shark- references.com, World Wide Web electronic publication, Version 08/2011; ISSN: 2195-6499 © Edited By: Jürgen Pollerspöck, Benediktinerring 34, D-94569 Stephansposching; Germany Please support www.shark-references.com Please send me missing, not listed references! Send me publications that are not incorporated so far (marked in red lettering)! - 2 - please inform me about missing papers E-Mail: [email protected] www.shark-references.com -

Middle-Late Triassic Chondrichthyans Remains from the Betic Range (Spain)
J Iber Geol DOI 10.1007/s41513-017-0027-1 RESEARCH PAPER Middle-Late Triassic chondrichthyans remains from the Betic Range (Spain) 1,2 1 2 2 E. Manzanares • C. Pla • H. G. Ferro´n • H. Botella Received: 14 March 2017 / Accepted: 5 July 2017 Ó Springer International Publishing AG 2017 Abstract recorded in Middle-Upper Triassic rocks from the Betic Purpose In the present study, we described, for first time, Domain could be used as nursery areas. the chondrichthyan fauna from several Middle-Late Tri- assic sections in the Betic Domain and compare it with Keywords Chondricthyans Á Ladinian Á Carnian Á other recent described coeval faunas from the Iberian Palaeocommunities Á Betic Range Ranges. Methods Specimens were retrieved after the dissolution Resumen (with 10% acetic acid) of carbonate rocks. Objetivo En el presente estudio describimos por primera Results The assemblage comprises of seven species vez, la fauna de condrictios de diversas secciones del belonging to six genera (Hybodus plicatilis, Omanoselache Tria´sico Medio-Tardı´o en el Dominio Be´tico y lo compa- bucheri comb. nov., Omanoselache contrarius comb. nov., ramos con otras faunas parecidas descritas recientemente Lonchidion derenzii, Lissodus aff. L. lepagei, Pseudo- en la Cordillera Ibe´rica. dalatias henarejensis and cf. Rhomaleodus budurovi), most Metodologı´a Los ejemplares fueron recuperados despue´s of them non-nesoselachian. Chondrichthyans remains de la disolucio´n (con a´cido ace´tico al 10%) de rocas occur in levels dating from Ladinian to Carnian according carbonatadas. with bivalves, ammonoids and conodonts. Resultados La asociacio´n se compone de siete especies Conclusions The findings are comparable, in taxonomical pertenecientes a seis ge´neros (Hybodus plicatilis, Omano- terms, to the chondrichthyan fauna from the Ladinian of selache bucheri comb. -

Family-Group Names of Fossil Fishes
European Journal of Taxonomy 466: 1–167 ISSN 2118-9773 https://doi.org/10.5852/ejt.2018.466 www.europeanjournaloftaxonomy.eu 2018 · Van der Laan R. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:1F74D019-D13C-426F-835A-24A9A1126C55 Family-group names of fossil fishes Richard VAN DER LAAN Grasmeent 80, 1357JJ Almere, The Netherlands. Email: [email protected] urn:lsid:zoobank.org:author:55EA63EE-63FD-49E6-A216-A6D2BEB91B82 Abstract. The family-group names of animals (superfamily, family, subfamily, supertribe, tribe and subtribe) are regulated by the International Code of Zoological Nomenclature. Particularly, the family names are very important, because they are among the most widely used of all technical animal names. A uniform name and spelling are essential for the location of information. To facilitate this, a list of family- group names for fossil fishes has been compiled. I use the concept ‘Fishes’ in the usual sense, i.e., starting with the Agnatha up to the †Osteolepidiformes. All the family-group names proposed for fossil fishes found to date are listed, together with their author(s) and year of publication. The main goal of the list is to contribute to the usage of the correct family-group names for fossil fishes with a uniform spelling and to list the author(s) and date of those names. No valid family-group name description could be located for the following family-group names currently in usage: †Brindabellaspidae, †Diabolepididae, †Dorsetichthyidae, †Erichalcidae, †Holodipteridae, †Kentuckiidae, †Lepidaspididae, †Loganelliidae and †Pituriaspididae. Keywords. Nomenclature, ICZN, Vertebrata, Agnatha, Gnathostomata. -

Progressive Palaeontology 2007
Progressive Palaeontology 2007 Thursday 12th – Saturday 14th April Department of Earth Sciences, University of Bristol 2 Welcome and acknowledgements We are pleased to welcome you to the Department of Earth Sciences at the University of Bristol for this year’s Progressive Palaeontology meeting. This appears to be the best attended and most presentation-packed Progressive Palaeontology on record – we are just shy of 60 delegates and there are 30 presentations in total (24 oral and 6 poster). As always Progressive Palaeontology’s aim is to provide a framework in which to present ideas and discuss work with those at a similar stage in their career. In order to achieve this the Friday presentation session is supplemented by two social events and a field trip. On Thursday evening there is a pre-conference gathering at the Berkeley pub (opposite the department on Queen’s Road) and following the presentation sessions on Friday there will also be an evening reception/dinner at Bristol Zoo. This year’s field trip will be to Aust Cliff (a site famous for its Late Triassic fish fossils – see the brief guide at the end of this booklet). We are lucky to have two experts on this site joining us on the day and we have scheduled a reasonably late start so please do come along if you have registered! We would also like to take this opportunity to thank the following individuals who have helped in supporting, preparing and running this year’s meeting: The Palaeontological Association and the University of Bristol Alumni Foundation for providing sponsorship. -

2. Fossil Record and Origin of Squaliform Sharks Jürgen Kriwet* Staatliches Museum Für Naturkunde, Rosenstein 1, 70191 Stuttgart, Germany
Biology and Management of Dogfish Sharks, 19–38 © 2009 by the American Fisheries Society 2. Fossil Record and Origin of Squaliform Sharks Jürgen Kriwet* Staatliches Museum für Naturkunde, Rosenstein 1, 70191 Stuttgart, Germany Stefanie Klug Museum für Naturkunde, Humboldt-Universität zu Berlin, Invalidenstr. 43, D-10115, Berlin, Germany Abstract.—Squaliform sharks constitute a monophyletic group of predominantly deep-water neose- lachians. Their fossil record mainly consists of isolated teeth; complete skeletons or skeletal remains are very rare. The quality of the fossil record of squaliform sharks is analyzed using a phylogenetic hypothesis based on a supertree to establish the timing of cladogenetic events, those related to descent from a common ancestor, and gaps in the fossil record. The supertree is the most inclusive estimate of squaliform interrelationships that has been proposed to date and contains 23 fossil and extant members of all major groups. In addition, the simple completeness metric is used to examine the quality of the fossil record of squaliforms as an independent measure. Although different (48% and 61%, respectively), both measures indicate that the fossil record of squaliforms is very incomplete considering that most living and extinct squaliforms are deep-water sharks and corresponding sediments are very scarce. Gaps in the fossil record range from 5 to 100 million years. The most basal and stratigraphically oldest group within Squaliformes consists of Squalus and †Protosqualus1. The phylogenetic hypothesis indicates a gap in the fossil record of Squalus spp. of about 25–30 million years. Our results show a postJurassic origination of squaliforms in the shallow waters of the northern Tethyal margin. -

Fascicolo III - Unità Validate
CARTA GEOLOGICA D’ITALIA 1:50.000 - CATALOGO DELLE FORMAZIONI 1 PRESIDENZA DEL CONSIGLIO DEI MINISTRI DIPARTIMENTO PER I SERVIZI TECNICI NAZIONALI SERVIZIO GEOLOGICO QUADERNI serie III Volume 7 CARTA GEOLOGICA D’ITALIA - 1:50.000 CATALOGO DELLE FORMAZIONI Fascicolo III - Unità validate a cura della COMMISSIONE ITALIANA DI STRATIGRAFIA DELLA SOCIETÀ GEOLOGICA ITALIANA ACCORDO DI PROGRAMMA SGN - CNR L. DELFRATI (1), P. FALORNI (2), G. GROPPELLI (3), F.M. PETTI (4) Impaginazione grafica P. I ZZO (4) (1) Dipartimento di Scienze della Terra “A. Desio”, Università degli Studi di Milano - [email protected] (2) Dipartimento di Scienze della Terra, Università degli Studi di Firenze - [email protected] (3) Istituto per la Dinamica dei Processi Ambientali, C.N.R., Sez. di Milano - [email protected] (4) c/o Dipartimento di Scienze della Terra, Università di Roma “La Sapienza” - [email protected] ROMA 2002 QUADERNI DEL SERVIZIO GEOLOGICO, SERIE III 1. Carta Geologica d’Italia - 1:50.000. Guida al rilevamento. 2. Carta Geologica d’Italia - 1:50.000. Guida alla rappresentazione dei dati. 3. Carta Geologica d’Italia - 1:50.000. Guida all’informatizzazione. 4. Carta Geomorfologica d’Italia - 1:50.000. Guida al rilevamento. 5. Carta Idrogeologica d’Italia - 1:50.000. Guida al rilevamento e alla rappresentazione. 6. Carta Geologica d’Italia - 1:50.000. Banca dati geologici (linee guida per l’informatizzazione e per l’allestimento per la stampa dalla banca dati). 7. Carta Geologica - 1:50.000. Catalogo delle Formazioni: - Fascicolo I - Unità validate. - Fascicolo II - Unità non validate (Unità da riclassificare e/o da abbandonare). -

The Rhaetian Vertebrates of Chipping Sodbury, South Gloucestershire
Proceedings of the Geologists’ Association 127 (2016) 40–52 Contents lists available at ScienceDirect Proceedings of the Geologists’ Association jo urnal homepage: www.elsevier.com/locate/pgeola The Rhaetian vertebrates of Chipping Sodbury, South Gloucestershire, UK, a comparative study a a,b,c a a, Rebecca J. Lakin , Christopher J. Duffin , Claudia Hildebrandt , Michael J. Benton * a School of Earth Sciences, University of Bristol, BS8 1RJ, UK b 146 Church Hill Road, Sutton, Surrey SM3 8NF, UK c Earth Sciences Department, The Natural History Museum, Cromwell Road, London SW7 5BD, UK A R T I C L E I N F O A B S T R A C T Article history: Microvertebrates are common in the basal bone bed of the Westbury Formation of England, Received 18 November 2015 documenting a fauna dominated by fishes that existed at the time of the Rhaetian Transgression, some Received in revised form 9 February 2016 206 Myr ago. Two sites near Chipping Sodbury, south Gloucestershire, Barnhill Quarry and Chipping Accepted 14 February 2016 Sodbury railway cutting, show differing faunas. Top predators are the large bony fish Severnichthys and Available online 22 March 2016 the shark Hybodus cloacinus, which preyed on smaller sharks such as Lissodus and Rhomphaiodon. These fishes in turn may have fed on a mixed diet of other fishes and invertebrates, and Lissodus was a shell Keywords: crusher. Comparisons of these faunas with others described recently from the Bristol region, and from Late Triassic Devon, indicate remarkable faunal similarities in the Rhaetian basal Westbury Formation bone bed over Systematics Chondrichthyes a wide area, based on a variety of ecological statistics that document species diversities and relative Actinopterygii abundances.