Intracranial Stimulation of the Trigeminal Nerve in Man II
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Blink Reflex, H-Reflex and Nerve-Conduction Alterations In
Lepr Rev (2006) 77, 114–120 Blink reflex, H-reflex and nerve-conduction alterations in leprosy patients ANA BERTHA MORA-BRAMBILA*, BENJAMI´N TRUJILLO-HERNA´ NDEZ**, RAFAEL COLL-CARDENAS***, MIGUEL HUERTA***, XO´ CHITL TRUJILLO***, CLEMENTE VA´ SQUEZ***, BERTHA ALICIA OLMEDO-BUENROSTRO*, REBECA O. MILLAN-GUERRERO** & ALEJANDRO ELIZALDE*** *Facultad de Enfermerı´a, Universidad de Colima, Colima, Me´xico **Unidad de Investigacio´n en Epidemiologı´a Clı´nica, Hospital General de Zona y Medicina Familiar No. 1, Instituto Mexicano del Seguro Social, Colima, Colima, Me´xico ***Centro Universitario de Investigaciones Biome´dicas, Universidad de Colima, Colima, Me´xico Accepted for publication 14 February 2006 Summary Peripheral nerve lesions are the most important cause of disability in leprosy patients. Electrophysiological studies are used in the diagnosis and prognosis of neuropathy. Nerve conduction is the most frequently used electrophysiological test method to detect neuropathy, although it evaluates only a part of the peripheral nervous system. Blink reflex and H-reflex are electrophysiological tests which evaluate facial and trigeminal nerve function. This study determined the frequencies of blink reflex, H-reflex and motor and sensory nerve conduction alterations in twenty five heterogeneous, clinic patients with lepromatous leprosy and a control group of 20 healthy subjects. Study results showed a decrease in motor and sensory nerve conduction in 40% and 30%, respectively. In blink reflex (BR), right R1 was altered in latency. in 20% of patients, left R1 in 20%, right ipsilateral R2 in 16%, left ipsilateral R2 in 20%, and right and left contralateral R2 were altered in 32% of patients. There was an absence of H-reflex in 16% (n ¼ 4) and prolonged latency in 4% (n ¼ 1). -

Neurophysiological Aspects of the Trigeminal Sensory System: an Update
Rev. Neurosci. 2018; 29(2): 115–123 Frederic Van der Cruyssen* and Constantinus Politis Neurophysiological aspects of the trigeminal sensory system: an update https://doi.org/10.1515/revneuro-2017-0044 Keywords: infraorbital; mandibular; neurophysiology; Received June 21, 2017; accepted July 20, 2017; previously published ophthalmic nerve; oral somatosensory functioning; online November 8, 2017 trigeminal sensory system. Abstract: The trigeminal system is one of the most complex cranial nerve systems of the human body. Research on it has vastly grown in recent years and concentrated more and more on molecular mechanisms and pathophysiology, Introduction but thorough reviews on this topic are lacking, certainly Knowledge about physiological aspects of the trigeminal on the normal physiology of the trigeminal sensory system. system today is largely based on animal models (Akerman Here we review the current literature on neurophysiology and Goadsby, 2015; Herta et al., 2017), cadaver studies of the trigeminal nerve from peripheral receptors up to its (Ezure et al., 2001; Williams et al., 2003) or extrapola- central projections toward the somatosensory cortex. We tions from peripheral nerve functioning. Human studies focus on the most recent scientific discoveries and describe are frequently limited to pathophysiology and lack proper historical relevant research to substantiate further. One study designs (Tanaka and Zhao, 2016; Goadsby et al., chapter on new insights of the pathophysiology of pain 2017). Neurophysiological research in this area is difficult at the level of the trigeminal system is added. A database due to the invasive character of most neurophysiological search of Medline, Embase and Cochrane was conducted tests, the small caliber of fibers, high density of receptors, with the search terms ‘animal study’, ‘neurophysiology’, cross-connections between different cranial nerves, dif- ‘trigeminal’, ‘oral’ and ‘sensory’. -

Innervation of the Temporomandibular Joint Can Be Discussed It Is Necessary First to Describe Its Embryology, Gfoss Anatomy and Microscopic Appe¿Ìrance
à8.ì 'R? INNERVATION OF THE TEMPOROMAI\DIBULAR J AN EXPERIMENTAL AMMAL MODEL USING AUSTRALIAN MERINO STIEEP ABDOLGHAFAR TAHMASEBI-SARVESTANI' B. Sc, M. Sc Thesis submitted for the degree of DOCTOR OF PHILOSOPHY In The Department of Anatomical Sciences The University of Adelaide (Faculty of Medicine)' Adelaide, South Australia, 5005 April, L997 tfüs tñesisis [elicatelø nl wtfe Aggñleñ ø¡tlour g4.arzi"e tfr.re e c friûfren Ía fiera ñ, fo zic ñ atú fi l-1 ACKNOWLEDGMENTS I am greatly indebted to my supervisors Dr. Ray Tedman and Professor Alastair Goss who first inrroduced me to this freld of study and providing me with the opportunity to carry out this work. I wish to thank them for their constant interest and guidance throughout the course of this study. I am also indebted to the scholarship committee of the Shiraz Medical Science University and Ministry of Health and Medical Education, Iran for gânting me a 4 year scholarship to study at the Universiry of Adelaide. I thank professor Goss and the Japanese Surgical Research team for their expertise in surgical animal models, and Professor July Polak and Dr Mika Hukkanen, Royal postgraduate Medical School London University for their expertise in immunohistochemistry and for providing some of the antisera used in the neuropeptide studies. I would also like to thank Professor Ian Gibbins, Department of Anatomy and Histology of the Flinders Medical Centre for, without the use of his laboratories, materials, and expertise, the double and triple labelling parts of the immunocytochemical work would not have occurred. I also orwe many thanks to Susan Matthew, a senior laboratory officer for her skilful technical assistance in double and triple immunocytochemistry. -
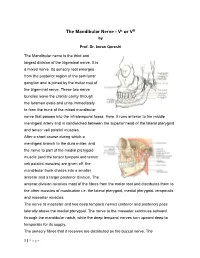
The Mandibular Nerve - Vc Or VIII by Prof
The Mandibular Nerve - Vc or VIII by Prof. Dr. Imran Qureshi The Mandibular nerve is the third and largest division of the trigeminal nerve. It is a mixed nerve. Its sensory root emerges from the posterior region of the semilunar ganglion and is joined by the motor root of the trigeminal nerve. These two nerve bundles leave the cranial cavity through the foramen ovale and unite immediately to form the trunk of the mixed mandibular nerve that passes into the infratemporal fossa. Here, it runs anterior to the middle meningeal artery and is sandwiched between the superior head of the lateral pterygoid and tensor veli palatini muscles. After a short course during which a meningeal branch to the dura mater, and the nerve to part of the medial pterygoid muscle (and the tensor tympani and tensor veli palatini muscles) are given off, the mandibular trunk divides into a smaller anterior and a larger posterior division. The anterior division receives most of the fibres from the motor root and distributes them to the other muscles of mastication i.e. the lateral pterygoid, medial pterygoid, temporalis and masseter muscles. The nerve to masseter and two deep temporal nerves (anterior and posterior) pass laterally above the medial pterygoid. The nerve to the masseter continues outward through the mandibular notch, while the deep temporal nerves turn upward deep to temporalis for its supply. The sensory fibres that it receives are distributed as the buccal nerve. The 1 | P a g e buccal nerve passes between the medial and lateral pterygoids and passes downward and forward to emerge from under cover of the masseter with the buccal artery. -

Morphometry and Morphology of Foramen Petrosum in Indian Population
Basic Sciences of Medicine 2020, 9(1): 8-9 DOI: 10.5923/j.medicine.20200901.02 Morphometry and Morphology of Foramen Petrosum in Indian Population Rajani Singh1,*, Nand Kishore Gupta1, Raj Kumar2 1Department of Anatomy, Uttar Pradesh University of Medical Sciences Saifai 206130 Etawah UP India 2Department of Neurosugery Uttar Pradesh University of Medical Sciences Saifai 206130 Etawah UP India Abstract Greater wing of sphenoid contains three constant foramina, Foramen ovale, foramen rotundum and foramen spinosum. The presence of foramen Vesalius and foramen petrosum are inconsistent. Normally foramen ovale transmits mandibular nerve, accessory meningeal artery, lesser petrosal nerve and emissary vein. When foramen petrosum is present, lesser petrosal nerve passes through petrosal foramen instead of foramen ovale. Lesser petrosal nerve distribute postganglionic fibers from otic ganglion to parotid gland. In absence of knowledge of petrosal foramen transmitting lesser petrosal nerve, the clinician may damage the nerve during skull base surgery creating complications like hyperemia of face and profuse salivation from the parotid gland (following atropine administration), lacrimation (crocodile tears syndrome) and mucus nasal secretion. Considering clinical implications associated with petrosal foramen, the study was carried out. The aim of the study is to determine the prevalence of petrosal foramen in Indian Population and to bring out associated clinical significance. The study was conducted in the department of Anatomy UPUMS Saifai Etawah Indian. 30 half skulls were observed for the presence of petrosal foramina and morphometry was also done. Literature search was carried out, our findings were compared with previous work and associated clinical implications were bought out. Keywords Petrosal foramen, Lesser petrosal nerve, Foramen ovale patients. -

Territorial and Extraterritorial Trigeminocardiac Reflex: a Review for the Neurosurgeon and a Type IV Reflex Vignette
Open Access Review Article DOI: 10.7759/cureus.11646 Territorial and Extraterritorial Trigeminocardiac Reflex: A Review for the Neurosurgeon and a Type IV Reflex Vignette Daniel S. Leon-Ariza 1 , Juan S. Leon-Ariza 2 , Mayra A. Gualdron 3 , Jaime Bayona-Prieto 4 , Fidias E. Leon- Sarmiento 5, 6, 7 1. School of Medicine, Santander University-UDES, Bucaramanga, COL 2. Neuroscience, Mediciencias Research Group, Miami, USA 3. Faculty of Medicine, Unicolsanitas, Bogota, COL 4. Cirineo Research Group, Unicolciencias, Bucaramanga, COL 5. Environmental Health, Florida International University, Miami, USA 6. Neurology, Baptist Health South Florida, Miami Neuroscience Institute, Miami, USA 7. Internal Medicine, National University, Bogota, COL Corresponding author: Fidias E. Leon-Sarmiento, [email protected] Abstract The trigeminocardiac reflex (TCR) is a complex and, sometimes, fatal event triggered by overstimulation of the trigeminal nerve (TN) and its territorial and spinal cord branches. We reviewed and compiled for the neurosurgeon key aspects of the TCR that include a novel and straightforward classification, as well as morphophysiology, pathophysiology, neuromonitoring and neuromodulation features. Further, we present intraoperative data from a patient who developed extraterritorial, or type IV, TCR while undergoing a cervical surgery. TCR complexity, severity and unwanted outcomes indicate that this event should not be underestimated or overlooked in the surgical room. Timely TCR recognition in surgical settings is valuable for applying effective intraoperative management to prevent catastrophic outcomes. Categories: Otolaryngology, Neurosurgery, Anatomy Keywords: trigeminocardiac reflex trigeminal nerve, spinal cord, neurophysiology, neuromonitoring, neuromodulation Introduction And Background The trigeminocardiac reflex (TCR) is a complex neurovascular reflex triggered by overstimulating the trigeminal nerve (TN) and its anastomosis. -

Brainstem Dysfunction in Critically Ill Patients
Benghanem et al. Critical Care (2020) 24:5 https://doi.org/10.1186/s13054-019-2718-9 REVIEW Open Access Brainstem dysfunction in critically ill patients Sarah Benghanem1,2 , Aurélien Mazeraud3,4, Eric Azabou5, Vibol Chhor6, Cassia Righy Shinotsuka7,8, Jan Claassen9, Benjamin Rohaut1,9,10† and Tarek Sharshar3,4*† Abstract The brainstem conveys sensory and motor inputs between the spinal cord and the brain, and contains nuclei of the cranial nerves. It controls the sleep-wake cycle and vital functions via the ascending reticular activating system and the autonomic nuclei, respectively. Brainstem dysfunction may lead to sensory and motor deficits, cranial nerve palsies, impairment of consciousness, dysautonomia, and respiratory failure. The brainstem is prone to various primary and secondary insults, resulting in acute or chronic dysfunction. Of particular importance for characterizing brainstem dysfunction and identifying the underlying etiology are a detailed clinical examination, MRI, neurophysiologic tests such as brainstem auditory evoked potentials, and an analysis of the cerebrospinal fluid. Detection of brainstem dysfunction is challenging but of utmost importance in comatose and deeply sedated patients both to guide therapy and to support outcome prediction. In the present review, we summarize the neuroanatomy, clinical syndromes, and diagnostic techniques of critical illness-associated brainstem dysfunction for the critical care setting. Keywords: Brainstem dysfunction, Brain injured patients, Intensive care unit, Sedation, Brainstem -

The Neurological Exam
The Neurological Exam Introduction to the Neurological Exam The neurological exam consists of the following components: 1. Higher cognitive function as assessed by the mental status examination. (This will be addressed elsewhere in the course.) 2. Cranial nerves 3. Motor system 4. Sensory systems 5. Stance and gait I Olfactory Nerve Examination Technique: stimulant should be non-irritating test one nostril at a time with the opposite side occluded patient should not be able to see the stimulus cloves ideal stimulant since it preserves it’s scent improvise at bedside with soap, toothpaste, or perfume Normal Response: to perceive the scent with either nostril Abnormal Response: a unilateral loss is more likely to be significant and may imply a structural brain lesion affecting the olfactory bulb or tract, but could also be due to local causes such as a deviated septum or blocked nasal passage bilateral loss can occur with rhinitis or damage to the cribriform plate II Optic Nerve - Visual Acuity Examination Technique: each eye is tested separately. test best corrected vision using eyeglasses. any patient with uncorrected visual acuity of less than 20/20 should be examined with a pinhole. Improvement of vision through a pinhole indicates that the error is refractive. test distant vision using a Snellen chart at 10 or 20 feet. II Optic Nerve - Visual Fields A. Peripheral visual field (a) wiggling fingers (b) counting fingers (c) white pin B. Central visual field (a) red pin Examination Technique: visual fields are assessed by confrontation , i.e. the examiner compares the patient’s visual field to their own and assumes that theirs is normal. -

Clinical Anatomy of the Trigeminal Nerve
Clinical Anatomy of Trigeminal through the superior orbital fissure Nerve and courses within the lateral wall of the cavernous sinus on its way The trigeminal nerve is the fifth of to the trigeminal ganglion. the twelve cranial nerves. Often Ophthalmic Nerve is formed by the referred to as "the great sensory union of the frontal nerve, nerve of the head and neck", it is nasociliary nerve, and lacrimal named for its three major sensory nerve. Branches of the ophthalmic branches. The ophthalmic nerve nerve convey sensory information (V1), maxillary nerve (V2), and from the skin of the forehead, mandibular nerve (V3) are literally upper eyelids, and lateral aspects "three twins" carrying information of the nose. about light touch, temperature, • The maxillary nerve (V2) pain, and proprioception from the enters the middle cranial fossa face and scalp to the brainstem. through foramen rotundum and may or may not pass through the • The three branches converge on cavernous sinus en route to the the trigeminal ganglion (also called trigeminal ganglion. Branches of the semilunar ganglion or the maxillary nerve convey sensory gasserian ganglion), which contains information from the lower eyelids, the cell bodies of incoming sensory zygomae, and upper lip. It is nerve fibers. The trigeminal formed by the union of the ganglion is analogous to the dorsal zygomatic nerve and infraorbital root ganglia of the spinal cord, nerve. which contain the cell bodies of • The mandibular nerve (V3) incoming sensory fibers from the enters the middle cranial fossa rest of the body. through foramen ovale, coursing • From the trigeminal ganglion, a directly into the trigeminal single large sensory root enters the ganglion. -

Computed Tomography of the Buccomasseteric Region: 1
605 Computed Tomography of the Buccomasseteric Region: 1. Anatomy Ira F. Braun 1 The differential diagnosis to consider in a patient presenting with a buccomasseteric James C. Hoffman, Jr. 1 region mass is rather lengthy. Precise preoperative localization of the mass and a determination of its extent and, it is hoped, histology will provide a most useful guide to the head and neck surgeon operating in this anatomically complex region. Part 1 of this article describes the computed tomographic anatomy of this region, while part 2 discusses pathologic changes. The clinical value of computed tomography as an imaging method for this region is emphasized. The differential diagnosis to consider in a patient with a mass in the buccomas seteric region, which may either be developmental, inflammatory, or neoplastic, comprises a rather lengthy list. The anatomic complexity of this region, defined arbitrarily by the soft tissue and bony structures including and surrounding the masseter muscle, excluding the parotid gland, makes the accurate anatomic diagnosis of masses in this region imperative if severe functional and cosmetic defects or even death are to be avoided during treatment. An initial crucial clinical pathoanatomic distinction is to classify the mass as extra- or intraparotid. Batsakis [1] recommends that every mass localized to the cheek region be considered a parotid tumor until proven otherwise. Precise clinical localization, however, is often exceedingly difficult. Obviously, further diagnosis and subsequent therapy is greatly facilitated once this differentiation is made. Computed tomography (CT), with its superior spatial and contrast resolution, has been shown to be an effective imaging method for the evaluation of disorders of the head and neck. -

Atlas of the Facial Nerve and Related Structures
Rhoton Yoshioka Atlas of the Facial Nerve Unique Atlas Opens Window and Related Structures Into Facial Nerve Anatomy… Atlas of the Facial Nerve and Related Structures and Related Nerve Facial of the Atlas “His meticulous methods of anatomical dissection and microsurgical techniques helped transform the primitive specialty of neurosurgery into the magnificent surgical discipline that it is today.”— Nobutaka Yoshioka American Association of Neurological Surgeons. Albert L. Rhoton, Jr. Nobutaka Yoshioka, MD, PhD and Albert L. Rhoton, Jr., MD have created an anatomical atlas of astounding precision. An unparalleled teaching tool, this atlas opens a unique window into the anatomical intricacies of complex facial nerves and related structures. An internationally renowned author, educator, brain anatomist, and neurosurgeon, Dr. Rhoton is regarded by colleagues as one of the fathers of modern microscopic neurosurgery. Dr. Yoshioka, an esteemed craniofacial reconstructive surgeon in Japan, mastered this precise dissection technique while undertaking a fellowship at Dr. Rhoton’s microanatomy lab, writing in the preface that within such precision images lies potential for surgical innovation. Special Features • Exquisite color photographs, prepared from carefully dissected latex injected cadavers, reveal anatomy layer by layer with remarkable detail and clarity • An added highlight, 3-D versions of these extraordinary images, are available online in the Thieme MediaCenter • Major sections include intracranial region and skull, upper facial and midfacial region, and lower facial and posterolateral neck region Organized by region, each layered dissection elucidates specific nerves and structures with pinpoint accuracy, providing the clinician with in-depth anatomical insights. Precise clinical explanations accompany each photograph. In tandem, the images and text provide an excellent foundation for understanding the nerves and structures impacted by neurosurgical-related pathologies as well as other conditions and injuries. -

Physiologic Factors for Dental Anesthesia Injections
ARE YOU NUMB YET? THE ANATOMY OF LOCAL ANESTHESIA PART 2: TECHNIQUES PHYSIOLOGIC FACTORS FOR DENTAL ANESTHESIA Alan W. Budenz, MS, DDS, MBA INJECTIONS Dept. of Biomedical Sciences and Vice Chair of Diagnostic Sciences & Services, Dept. of Dental Practice University of the Pacific, Arthur A. Dugoni School of Dentistry San Francisco, California Success versus Failure [email protected] Failed Anesthetic: Measuring the Problem Physiology of Anesthetic Agents One of every three patients is not properly numb when the dentist or hygienist is ready to start (or actually starts) a dental procedure. How do we assess anesthesia? Is this “failed anesthetic”? 60% Question the patient Soft tissue only 50% * Probe the area 46% Average 40% 42% 41% Failure 38% Rate is Cold test 30% 29% Pulpal tissue 31% Electric pulp tester 20% 19% 20% 17% 15% How is anesthetic success defined in studies? 10% Frequency Frequency Anesthetic Failedof Ideal: 2 consecutive 80/80 readings with EPT within 15 0% IAN Blocks - 15 min. after injection Maxillary infiltrations - 10 min. after injection minutes of injection (and sustained for 60 mins) Delayed pulpal onset: occurs in the mandible of 19 – 27% Slide courtesy Dr. Mic Falkel of patients (even though soft tissue is numb) Delayed over 30 minutes in 8% Nusstein J et al. The challenges of successful * Average failure rate reported across 38 published studies mandibular anesthesia, Inside Dentistry, May 2008 Physiology of Anesthetic Agents Blocks versus Infiltrations Onset of anesthesia: Advantages of infiltrations 1. Dependent upon anesthetic agent 1. Faster onset Concentration 2. Diffusion to the site Simple Lipid solubility 3.