Polyploidy and Ecological Adaptation in Wild Yarrow
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Notas Sobre El Género Ambrosia (Asteraceae: Ambrosiinae) En Chile Notes on the Genus Ambrosia (Asteraceae: Ambrosiinae) in Chile
Chloris Chilensis 23 (1): 77-83. 2020. NOTAS SOBRE EL GÉNERO AMBROSIA (ASTERACEAE: AMBROSIINAE) EN CHILE NOTES ON THE GENUS AMBROSIA (ASTERACEAE: AMBROSIINAE) IN CHILE Federico Luebert & Nicolás García Herbario EIF, Departamento de Silvicultura y Conservación de la Naturaleza, Universidad de Chile, Santiago, Chile E-mail: [email protected] RESUMEN Se discuten aspectos nomenclaturales y taxonómicos de tres especies de Ambrosia de Chile. Se concluye que el nombre Ambrosia artemisiifolia L. debe ser usado en lugar de A. elatior L.; el nombre A. cumanensis Kunth, en lugar de A. peruviana Willd.; y que el nombre A. tarapacana Phil., debe agregarse al catálogo de la flora de Chile hasta que nuevos estudios se hagan disponibles. El género, así considerado, queda representado en el país por siete especies. Se proporciona una clave para la determinación de las especies de Ambrosia presentes en Chile. Palabras clave: Compositae, nomenclatura, taxonomía. ABSTRACT Nomenclatural and taxonomic aspects of three Ambrosia species present in Chile are discussed. We conclude that the name Ambrosia artemisiifolia L. should be used instead of A. elatior L., the name A. cumanensis Kunth instead of A. peruviana Willd., and the name A. tarapacana Phil. should be added to the catalogue of the Chilean flora until new studies become available. The genus is thus represented in Chile by seven species. A key for the determination of the Chilean species of Ambrosia is provided. Key words: Compositae, Nomenclature, Taxonomy. Luebert & García: Ambrosia en Chile. Chloris Chilensis 23 (1): 77-83. 2020. INTRODUCCIÓN El género Ambrosia L. (Asteraceae) fue creado por Linneo para incluir cuatro especies (Linnaeus, 1753). -

Pollens - Weeds and Garden Plants, Alfalfa Medicago Sativa
Product: 1057 - Pollens - Weeds and Garden Plants, Alfalfa Medicago sativa Manufacturers of this Product Antigen Laboratories, Inc. - Liberty, MO (Lic. No. 468, STN No. 102223) Hollister-Stier Labs, LLC - Spokane, WA (Lic. No. 1272, STN No. 103888) ALK-Abello Inc. - Port Washington, NY (Lic. No. 1256, STN No. 103753) Nelco Laboratories, Inc. - Deer Park, NY (Lic. No. 459, STN No. 102192) Search Strategy PubMed: alfalfa allergen; allfalfa allergy; alfalfa immunotherapy; Medicago sativa allergy; Medicago sativa allergen; Medicago sativa immunotherapy; Alfalfa pollen Google: alfalfa allergy; alfalfa allergy adverse; alfalfa allergen; alfalfa allergen adverse; same search results performed for Medicago sativa Nomenclature According to ITIS, the scientific name is Medicago sativa (TSN 183623). The common name is alfalfa. The scientific and common names are correct and current. There are two subspecies listed: M. sativa ssp. falcate and sativa. The Medicago genus is found in the Fabaceae family. Parent Product 1057 - Pollens - Weeds and Garden Plants, Alfalfa Medicago sativa Published Data PMID: 9109711: One article described skin prick testing with inhalent allergens, including Alfalfa, Medicago sativa from "Hollister-Stier, (now Miles, Inc., Connecticut, USA)" on a Saudi Arabian population, including North American expatriots. PMID: 6733890: Positive skin test results were recorded for M. sativa (13.5%) in this Mediterranean study of pollen counts and pollinosis. Documents PubMed Number Author Year Published 563_ Ann Allergy Asthma Immunol-78-415.pdf 9109711 Suliaman, F.A. 1997 1090_ ClinicalAllergy14-249-258.pdf 6733890 Bousquet, J. 1984 Panels: Original Panel Recommendation Reclassification Panel Recommendation Diagnosis: none none Therapy: none none Page 1 of 218 9/9/2011 Product: 1058 - Pollens - Weeds and Garden Plants, Allscale Atriplex polycarpa Recommended Common Name Saltbush, Desert Manufacturers of this Product Antigen Laboratories, Inc. -

Baja California, Mexico, and a Vegetation Map of Colonet Mesa Alan B
Aliso: A Journal of Systematic and Evolutionary Botany Volume 29 | Issue 1 Article 4 2011 Plants of the Colonet Region, Baja California, Mexico, and a Vegetation Map of Colonet Mesa Alan B. Harper Terra Peninsular, Coronado, California Sula Vanderplank Rancho Santa Ana Botanic Garden, Claremont, California Mark Dodero Recon Environmental Inc., San Diego, California Sergio Mata Terra Peninsular, Coronado, California Jorge Ochoa Long Beach City College, Long Beach, California Follow this and additional works at: http://scholarship.claremont.edu/aliso Part of the Biodiversity Commons, Botany Commons, and the Ecology and Evolutionary Biology Commons Recommended Citation Harper, Alan B.; Vanderplank, Sula; Dodero, Mark; Mata, Sergio; and Ochoa, Jorge (2011) "Plants of the Colonet Region, Baja California, Mexico, and a Vegetation Map of Colonet Mesa," Aliso: A Journal of Systematic and Evolutionary Botany: Vol. 29: Iss. 1, Article 4. Available at: http://scholarship.claremont.edu/aliso/vol29/iss1/4 Aliso, 29(1), pp. 25–42 ’ 2011, Rancho Santa Ana Botanic Garden PLANTS OF THE COLONET REGION, BAJA CALIFORNIA, MEXICO, AND A VEGETATION MAPOF COLONET MESA ALAN B. HARPER,1 SULA VANDERPLANK,2 MARK DODERO,3 SERGIO MATA,1 AND JORGE OCHOA4 1Terra Peninsular, A.C., PMB 189003, Suite 88, Coronado, California 92178, USA ([email protected]); 2Rancho Santa Ana Botanic Garden, 1500 North College Avenue, Claremont, California 91711, USA; 3Recon Environmental Inc., 1927 Fifth Avenue, San Diego, California 92101, USA; 4Long Beach City College, 1305 East Pacific Coast Highway, Long Beach, California 90806, USA ABSTRACT The Colonet region is located at the southern end of the California Floristic Province, in an area known to have the highest plant diversity in Baja California. -

Vascular Plants of Humboldt Bay's Dunes and Wetlands Published by U.S
Vascular Plants of Humboldt Bay's Dunes and Wetlands Published by U.S. Fish and Wildlife Service G. Leppig and A. Pickart and California Department of Fish Game Release 4.0 June 2014* www.fws.gov/refuge/humboldt_bay/ Habitat- Habitat - Occurs on Species Status Occurs within Synonyms Common name specific broad Lanphere- Jepson Manual (2012) (see codes at end) refuge (see codes at end) (see codes at end) Ma-le'l Units UD PW EW Adoxaceae Sambucus racemosa L. red elderberry RF, CDF, FS X X N X X Aizoaceae Carpobrotus chilensis (Molina) sea fig DM X E X X N.E. Br. Carpobrotus edulis ( L.) N.E. Br. Iceplant DM X E, I X Alismataceae lanceleaf water Alisma lanceolatum With. FM X E plantain northern water Alisma triviale Pursh FM X N plantain Alliaceae three-cornered Allium triquetrum L. FS, FM, DM X X E leek Allium unifolium Kellogg one-leaf onion CDF X N X X Amaryllidaceae Amaryllis belladonna L. belladonna lily DS, AW X X E Narcissus pseudonarcissus L. daffodil AW, DS, SW X X E X Anacardiaceae Toxicodendron diversilobum Torrey poison oak CDF, RF X X N X X & A. Gray (E. Greene) Apiaceae Angelica lucida L. seacoast angelica BM X X N, C X X Anthriscus caucalis M. Bieb bur chevril DM X E Cicuta douglasii (DC.) J. Coulter & western water FM X N Rose hemlock Conium maculatum L. poison hemlock RF, AW X I X Daucus carota L. Queen Anne's lace AW, DM X X I X American wild Daucus pusillus Michaux DM, SW X X N X X carrot Foeniculum vulgare Miller sweet fennel AW, FM, SW X X I X Glehnia littoralis (A. -

Vascular Plants of Santa Cruz County, California
ANNOTATED CHECKLIST of the VASCULAR PLANTS of SANTA CRUZ COUNTY, CALIFORNIA SECOND EDITION Dylan Neubauer Artwork by Tim Hyland & Maps by Ben Pease CALIFORNIA NATIVE PLANT SOCIETY, SANTA CRUZ COUNTY CHAPTER Copyright © 2013 by Dylan Neubauer All rights reserved. No part of this publication may be reproduced without written permission from the author. Design & Production by Dylan Neubauer Artwork by Tim Hyland Maps by Ben Pease, Pease Press Cartography (peasepress.com) Cover photos (Eschscholzia californica & Big Willow Gulch, Swanton) by Dylan Neubauer California Native Plant Society Santa Cruz County Chapter P.O. Box 1622 Santa Cruz, CA 95061 To order, please go to www.cruzcps.org For other correspondence, write to Dylan Neubauer [email protected] ISBN: 978-0-615-85493-9 Printed on recycled paper by Community Printers, Santa Cruz, CA For Tim Forsell, who appreciates the tiny ones ... Nobody sees a flower, really— it is so small— we haven’t time, and to see takes time, like to have a friend takes time. —GEORGIA O’KEEFFE CONTENTS ~ u Acknowledgments / 1 u Santa Cruz County Map / 2–3 u Introduction / 4 u Checklist Conventions / 8 u Floristic Regions Map / 12 u Checklist Format, Checklist Symbols, & Region Codes / 13 u Checklist Lycophytes / 14 Ferns / 14 Gymnosperms / 15 Nymphaeales / 16 Magnoliids / 16 Ceratophyllales / 16 Eudicots / 16 Monocots / 61 u Appendices 1. Listed Taxa / 76 2. Endemic Taxa / 78 3. Taxa Extirpated in County / 79 4. Taxa Not Currently Recognized / 80 5. Undescribed Taxa / 82 6. Most Invasive Non-native Taxa / 83 7. Rejected Taxa / 84 8. Notes / 86 u References / 152 u Index to Families & Genera / 154 u Floristic Regions Map with USGS Quad Overlay / 166 “True science teaches, above all, to doubt and be ignorant.” —MIGUEL DE UNAMUNO 1 ~ACKNOWLEDGMENTS ~ ANY THANKS TO THE GENEROUS DONORS without whom this publication would not M have been possible—and to the numerous individuals, organizations, insti- tutions, and agencies that so willingly gave of their time and expertise. -

Vegetation Classification for San Juan Island National Historical Park
National Park Service U.S. Department of the Interior Natural Resource Stewardship and Science San Juan Island National Historical Park Vegetation Classification and Mapping Project Report Natural Resource Report NPS/NCCN/NRR—2012/603 ON THE COVER Red fescue (Festuca rubra) grassland association at American Camp, San Juan Island National Historical Park. Photograph by: Joe Rocchio San Juan Island National Historical Park Vegetation Classification and Mapping Project Report Natural Resource Report NPS/NCCN/NRR—2012/603 F. Joseph Rocchio and Rex C. Crawford Natural Heritage Program Washington Department of Natural Resources 1111 Washington Street SE Olympia, Washington 98504-7014 Catharine Copass National Park Service North Coast and Cascades Network Olympic National Park 600 E. Park Ave. Port Angeles, Washington 98362 . December 2012 U.S. Department of the Interior National Park Service Natural Resource Stewardship and Science Fort Collins, Colorado The National Park Service, Natural Resource Stewardship and Science office in Fort Collins, Colorado, publishes a range of reports that address natural resource topics. These reports are of interest and applicability to a broad audience in the National Park Service and others in natural resource management, including scientists, conservation and environmental constituencies, and the public. The Natural Resource Report Series is used to disseminate high-priority, current natural resource management information with managerial application. The series targets a general, diverse audience, and may contain NPS policy considerations or address sensitive issues of management applicability. All manuscripts in the series receive the appropriate level of peer review to ensure that the information is scientifically credible, technically accurate, appropriately written for the intended audience, and designed and published in a professional manner. -

Checklist of the Vascular Plants of San Diego County 5Th Edition
cHeckliSt of tHe vaScUlaR PlaNtS of SaN DieGo coUNty 5th edition Pinus torreyana subsp. torreyana Downingia concolor var. brevior Thermopsis californica var. semota Pogogyne abramsii Hulsea californica Cylindropuntia fosbergii Dudleya brevifolia Chorizanthe orcuttiana Astragalus deanei by Jon P. Rebman and Michael G. Simpson San Diego Natural History Museum and San Diego State University examples of checklist taxa: SPecieS SPecieS iNfRaSPecieS iNfRaSPecieS NaMe aUtHoR RaNk & NaMe aUtHoR Eriodictyon trichocalyx A. Heller var. lanatum (Brand) Jepson {SD 135251} [E. t. subsp. l. (Brand) Munz] Hairy yerba Santa SyNoNyM SyMBol foR NoN-NATIVE, NATURaliZeD PlaNt *Erodium cicutarium (L.) Aiton {SD 122398} red-Stem Filaree/StorkSbill HeRBaRiUM SPeciMeN coMMoN DocUMeNTATION NaMe SyMBol foR PlaNt Not liSteD iN THE JEPSON MANUAL †Rhus aromatica Aiton var. simplicifolia (Greene) Conquist {SD 118139} Single-leaF SkunkbruSH SyMBol foR StRict eNDeMic TO SaN DieGo coUNty §§Dudleya brevifolia (Moran) Moran {SD 130030} SHort-leaF dudleya [D. blochmaniae (Eastw.) Moran subsp. brevifolia Moran] 1B.1 S1.1 G2t1 ce SyMBol foR NeaR eNDeMic TO SaN DieGo coUNty §Nolina interrata Gentry {SD 79876} deHeSa nolina 1B.1 S2 G2 ce eNviRoNMeNTAL liStiNG SyMBol foR MiSiDeNtifieD PlaNt, Not occURRiNG iN coUNty (Note: this symbol used in appendix 1 only.) ?Cirsium brevistylum Cronq. indian tHiStle i checklist of the vascular plants of san Diego county 5th edition by Jon p. rebman and Michael g. simpson san Diego natural history Museum and san Diego state university publication of: san Diego natural history Museum san Diego, california ii Copyright © 2014 by Jon P. Rebman and Michael G. Simpson Fifth edition 2014. isBn 0-918969-08-5 Copyright © 2006 by Jon P. -

Plant Identification of Younger Lagoon Reserve
Plant Identification of Younger Lagoon Reserve A guide written by Rebecca Evans with help from Dr. Karen Holl, Elizabeth Howard, and Timothy Brown 1 Table of Contents Introduction to Plant Identification ............................................................................................. 3 Plant Index ................................................................................................................................. 6 Botanical Terminology ............................................................................................................. 12 Habits, Stem Conditions, Root Types ................................................................................ 12 Leaf Parts .......................................................................................................................... 13 Stem Features .................................................................................................................... 14 Leaf Arrangements ............................................................................................................ 16 Leaf Shape ........................................................................................................................ 18 Leaf Margins and Venation ............................................................................................... 20 Flowers and Inflorescences ................................................................................................ 21 Grasses ............................................................................................................................. -

Lanphere National Natural Landmark Evaluation
Evaluation of Lanphere / Ma-le’l Dunes Humboldt County, California For its Merit in Meeting National Significance Criteria as a National Natural Landmark to Represent Siskiyou – Klamath Coastal Sand Dunes in the North Pacific Border Biophysiographic Province Peter Alpert and James S. Kagan May 2019 Contact Information: Peter Alpert, Department of Biology, University of Massachusetts, Amherst, Massachusetts 01003 James S. Kagan, Oregon Biodiversity Information Center. Institute for Natural Resources – Portland, Portland State University. Mailstop: INR. P.O. Box 751, Portland, Oregon 97207. Citation: Alpert, P. and J.S. Kagan, 2019. Evaluation of Lanphere / Ma-le’l Dunes, Humboldt County, California for its Merit in Meeting National Significance Criteria as a National Natural Landmark to Represent Siskiyou – Klamath Coastal Sand Dunes in the North Pacific Border Biophysiographic Province. National Park Service Technical Report, National Natural Landmarks Program, Fort Collins, CO. Acknowledgements The authors would like to acknowledge the work of the many people who participated in the evaluation of the North Pacific Border province coastal dunes theme studies. The authors would also like to acknowledge the help, support and patience of our partners at the National Park Service, in particular Laurette Jenkins, who took this on as her first project, and has done a wonderful job, helping us with formatting, questions, and direction on the project. We also especially thank Steve Gibbons, who recently left the Pacific West Region’s NNL program job after 22 years to become the superintendent of Mount Lassen National Park and who inspired us to take on this report. Both authors would like to recognize Andrea Pickart, for her help, great photos, guidance, and advice. -

Plant Check List
1 Vascular Plants of MacKerricher State Park A Latin Binomial Common Name Glass Bch. only FERNS Athyriaceae Athyrium filix-femina var. cyclosorum lady fern Azollaceae Azolla filiculoides mosquito fern Blechnaceae Struthiopteris spicant deer fern Woodwardia fimbriata giant chain fern Cystopteridaceae Cystopteris fragilis fragile fern Dennstaedtiaceae Pteridium aquilinum var. pubescens bracken Dryopteridaceae Drypoteris arguta wood fern Polystichum munitum western sword fern Equisetaceae Equisetum arvense field horsetail Equisetum hyemale ssp. affine common scouring-rush Equisetum laevigatum smooth scouring-rush Equisetum telmateia ssp. braunii giant horsetail Ophioglossaceae Sceptridium multifidum moonwort; grape fern Polypodiaceae Polypodium californicum California polypody Polypdium calirhiza polypody fern Polypodium scouleri Pteridaceae Pentagramma triangularis goldback fern GYMNOSPERMS Cupressaceae Hesperocyparis macrocarpa* Monterey cypress Sequoia sempervirens coast redwood Pinaceae Abies grandis grand fir Pinus contorta ssp. contorta shore pine; beach pine Pinus muricata Bishop pine Pinus radiata* Monterey pine Pseudotsuga menziesii Douglas-fir NYMPHAEALES Nymphaeaceae Nuphar polysepala yellow pond-lily EUDICOTS Adoxaceae Sambucus racemosa var. racemosa red elderberry Aizoaceae Carpobrotus chilensis* sea fig; iceplant Carpobrotus edulis* Hottentot-fig; iceplant Lampranthus spectabilis* redflush iceplant Tetragonia tetragonioides* New Zealand spinach Anacardiaceae Toxicodendrom diversilobum poison-oak Apiaceae Angelica hendersonii -

SF Flora Sutro Baths V2014-04-06.Xlsx
ANNOTATED CHECKLIST OF THE VASCULAR PLANTS OF SUTRO BATHS (GGNRA) h i Habit/Habitats k c Major Group [Synonymy] / notes j f g Family Common Name rarity statusd b notes Period Scientific Name invasive statuse Flowering Flowering life form life duration Native: SF / CA # SF Occurrences # SF PTERIDOPHYTES - Spore-Bearing Ferns and Fern Allies Equisetaceae - Horsetail Family Rhizomatous. crk, Equisetum telmateia ssp. braunii giant horsetail fp 9n/aY/Y lak/pnd, mrsh/sp (mst) GYMNOSPERMS - Cone-bearing plants Cupressaceae - Cypress Family Hesperocyparis macrocarpa Monterey cypress [Cupressus m.] CA native not indigenous to SF. t p Evergreen. 41 n/a N/Y EUDICOTS - Flowering, seed-bearing plants producing two cotyledons (seed leaves) Aizoaceae - Fig-Marigold or Iceplant Family Carpobrotus chilensis sea fig [Mesembryanthemum c.] Cal-IPC: Moderate. h p Creeping succulent. 13 all yr. N/N Carpobrotus edulis iceplant, Hottentot fig [Mesembryanthemum edule] Cal-IPC: High. h p Creeping succulent. 27 all yr. N/N Lampranthus sp. redflush Not in Howell, et al. Non-specific report from Sutro Baths. h p 1 N/N Tetragonia tetragonioides New Zealand spinach Cal-IPC: Limited. h a 13 Apr–Sep N/N Apiaceae - Carrot Family Foeniculum vulgare sweet fennel Cal-IPC: High; SFWMA: Priority Weed. h p 34 May–Sep N/N Araliaceae - Ginseng Family Leaves on juvenile stems <= 35 cm, palmately 3–5-lobed, lobes ± Hedera helix English Ivy acute; hair white, hair rays spreading. Cal-IPC: High; SFWMA: v p Woody. 23 Aug–Nov N/N Priority Weed. Asteraceae - Sunflower Family beach bur, silver Ambrosia chamissonis [Franseria c.] Includes A.c var. -
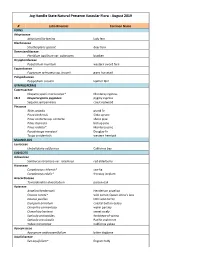
Plant Check List
Jug Handle State Natural Preserve Vascular Flora - August 2019 A1 Latin Binomial Common Name FERNS Athyriaceae Athyrium filix-femina lady fern Blechnaceae Struthiopteris spicant deer fern Dennstaedtiaceae Pteridium aqulinum var. pubescens bracken Dryopteridaceae Polystichum munitum western sword fern Equisetaceae Equisetum telmateia ssp. braunii giant horsetail Polypodiaceae Polypodium scouleri leather fern GYMNOSPERMS Cupressaceae Hesperocyparis macrocarpa* Monterey cypress 1B.2 Hesperocyparis pygmaea pygmy cypress Sequoia sempervirens coast redwood Pinaceae Abies grandis grand fir Picea sitchensis Sitka spruce Pinus contorta ssp. contorta shore pine Pinus muricata bishop pine Pinus radiata* Monterey pine Pseudotsuga menziesii Douglas-fir Tsuga occidentalis western hemlock MAGNOLIIDS Lauraceae Umbellularia californica California bay EUDICOTS Adoxaceae Sambucus racemosa var. racemosa red elderberry Aizoaceae Carpobrotus chilensis* sea-fig Carpobrotus edulis* freeway iceplant Anacardiaceae Toxicodendron diversilobum poison-oak Apiaceae Angelica hendersonii Henderson angelica Daucus carota* wild carrot; Queen Anne's lace Daucus pusillus little wild carrot Eryngium armatum coastal button-celery Oenanthe sarmentosa water parsley Osmorhiza berteroi sweet cicely Sanicula arctopoides footsteps-of-spring Sanicula crassicaulis Pacific snakeroot Yabea microcarpa California yabea Apocynaceae Apocynum androsaemifolium bitter dogbane Aquifoliaceae Ilex aquifolium* English holly Aristolochiaceae Asarum caudatum wild ginger Asteraceae Achillea millefolium