Angiosperm Phylogeny Flowering Plant Systematics
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -

Alphabetical Lists of the Vascular Plant Families with Their Phylogenetic
Colligo 2 (1) : 3-10 BOTANIQUE Alphabetical lists of the vascular plant families with their phylogenetic classification numbers Listes alphabétiques des familles de plantes vasculaires avec leurs numéros de classement phylogénétique FRÉDÉRIC DANET* *Mairie de Lyon, Espaces verts, Jardin botanique, Herbier, 69205 Lyon cedex 01, France - [email protected] Citation : Danet F., 2019. Alphabetical lists of the vascular plant families with their phylogenetic classification numbers. Colligo, 2(1) : 3- 10. https://perma.cc/2WFD-A2A7 KEY-WORDS Angiosperms family arrangement Summary: This paper provides, for herbarium cura- Gymnosperms Classification tors, the alphabetical lists of the recognized families Pteridophytes APG system in pteridophytes, gymnosperms and angiosperms Ferns PPG system with their phylogenetic classification numbers. Lycophytes phylogeny Herbarium MOTS-CLÉS Angiospermes rangement des familles Résumé : Cet article produit, pour les conservateurs Gymnospermes Classification d’herbier, les listes alphabétiques des familles recon- Ptéridophytes système APG nues pour les ptéridophytes, les gymnospermes et Fougères système PPG les angiospermes avec leurs numéros de classement Lycophytes phylogénie phylogénétique. Herbier Introduction These alphabetical lists have been established for the systems of A.-L de Jussieu, A.-P. de Can- The organization of herbarium collections con- dolle, Bentham & Hooker, etc. that are still used sists in arranging the specimens logically to in the management of historical herbaria find and reclassify them easily in the appro- whose original classification is voluntarily pre- priate storage units. In the vascular plant col- served. lections, commonly used methods are systema- Recent classification systems based on molecu- tic classification, alphabetical classification, or lar phylogenies have developed, and herbaria combinations of both. -

Lessons from 20 Years of Plant Genome Sequencing: an Unprecedented Resource in Need of More Diverse Representation
bioRxiv preprint doi: https://doi.org/10.1101/2021.05.31.446451; this version posted May 31, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Lessons from 20 years of plant genome sequencing: an unprecedented resource in need of more diverse representation Authors: Rose A. Marks1,2,3, Scott Hotaling4, Paul B. Frandsen5,6, and Robert VanBuren1,2 1. Department of Horticulture, Michigan State University, East Lansing, MI 48824, USA 2. Plant Resilience Institute, Michigan State University, East Lansing, MI 48824, USA 3. Department of Molecular and Cell Biology, University of Cape Town, Rondebosch 7701, South Africa 4. School of Biological Sciences, Washington State University, Pullman, WA, USA 5. Department of Plant and Wildlife Sciences, Brigham Young University, Provo, UT, USA 6. Data Science Lab, Smithsonian Institution, Washington, DC, USA Keywords: plants, embryophytes, genomics, colonialism, broadening participation Correspondence: Rose A. Marks, Department of Horticulture, Michigan State University, East Lansing, MI 48824, USA; Email: [email protected]; Phone: (603) 852-3190; ORCID iD: https://orcid.org/0000-0001-7102-5959 Abstract The field of plant genomics has grown rapidly in the past 20 years, leading to dramatic increases in both the quantity and quality of publicly available genomic resources. With an ever- expanding wealth of genomic data from an increasingly diverse set of taxa, unprecedented potential exists to better understand the evolution and genome biology of plants. -

Phylogenomic Approach
Toward the ultimate phylogeny of Magnoliaceae: phylogenomic approach Sangtae Kim*1, Suhyeon Park1, and Jongsun Park2 1 Sungshin University, Korea 2 InfoBoss Co., Korea Mr. Carl Ferris Miller Founder of Chollipo Arboretum in Korea Chollipo Arboretum Famous for its magnolia collection 2020. Annual Meeting of Magnolia Society International Cholliop Arboretum in Korea. April 13th~22th, 2020 http://WWW.Chollipo.org Sungshin University, Seoul, Korea Dr. Hans Nooteboom Dr. Liu Yu-Hu Twenty-one years ago... in 1998 The 1st International Symposium on the Family Magnoliaceae, Gwangzhow Dr. Hiroshi Azuma Mr. Richard Figlar Dr. Hans Nooteboom Dr. Qing-wen Zeng Dr. Weibang Sun Handsome young boy Dr. Yong-kang Sima Dr. Yu-wu Law Presented ITS study on Magnoliaceae - never published Ten years ago... in 2009 Presented nine cp genome region study (9.2 kbp) on Magnoliaceae – published in 2013 2015 1st International Sympodium on Neotropical Magnoliaceae Gadalajara, 2019 3rd International Sympodium and Workshop on Neotropical Magnoliaceae Asterales Dipsacales Apiales Why magnolia study is Aquifoliales Campanulids (Euasterids II) Garryales Gentianales Laminales Solanales Lamiids important in botany? Ericales Asterids (Euasterids I) Cornales Sapindales Malvales Brassicales Malvids Fagales (Eurosids II) • As a member of early-diverging Cucurbitales Rosales Fabales Zygophyllales Celestrales Fabids (Eurosid I) angiosperms, reconstruction of the Oxalidales Malpighiales Vitales Geraniales Myrtales Rosids phylogeny of Magnoliaceae will Saxifragales Caryphyllales -

Reconstructing the Basal Angiosperm Phylogeny: Evaluating Information Content of Mitochondrial Genes
55 (4) • November 2006: 837–856 Qiu & al. • Basal angiosperm phylogeny Reconstructing the basal angiosperm phylogeny: evaluating information content of mitochondrial genes Yin-Long Qiu1, Libo Li, Tory A. Hendry, Ruiqi Li, David W. Taylor, Michael J. Issa, Alexander J. Ronen, Mona L. Vekaria & Adam M. White 1Department of Ecology & Evolutionary Biology, The University Herbarium, University of Michigan, Ann Arbor, Michigan 48109-1048, U.S.A. [email protected] (author for correspondence). Three mitochondrial (atp1, matR, nad5), four chloroplast (atpB, matK, rbcL, rpoC2), and one nuclear (18S) genes from 162 seed plants, representing all major lineages of gymnosperms and angiosperms, were analyzed together in a supermatrix or in various partitions using likelihood and parsimony methods. The results show that Amborella + Nymphaeales together constitute the first diverging lineage of angiosperms, and that the topology of Amborella alone being sister to all other angiosperms likely represents a local long branch attrac- tion artifact. The monophyly of magnoliids, as well as sister relationships between Magnoliales and Laurales, and between Canellales and Piperales, are all strongly supported. The sister relationship to eudicots of Ceratophyllum is not strongly supported by this study; instead a placement of the genus with Chloranthaceae receives moderate support in the mitochondrial gene analyses. Relationships among magnoliids, monocots, and eudicots remain unresolved. Direct comparisons of analytic results from several data partitions with or without RNA editing sites show that in multigene analyses, RNA editing has no effect on well supported rela- tionships, but minor effect on weakly supported ones. Finally, comparisons of results from separate analyses of mitochondrial and chloroplast genes demonstrate that mitochondrial genes, with overall slower rates of sub- stitution than chloroplast genes, are informative phylogenetic markers, and are particularly suitable for resolv- ing deep relationships. -

Calibrated Chronograms, Fossils, Outgroup Relationships, and Root Priors: Re-Examining the Historical Biogeography of Geraniales
bs_bs_banner Biological Journal of the Linnean Society, 2014, 113, 29–49. With 4 figures Calibrated chronograms, fossils, outgroup relationships, and root priors: re-examining the historical biogeography of Geraniales KENNETH J. SYTSMA1,*, DANIEL SPALINK1 and BRENT BERGER2 1Department of Botany, University of Wisconsin, Madison, WI 53706, USA 2Department of Biological Sciences, St. John’s University, Queens, NY 11439, USA Received 26 November 2013; revised 23 February 2014; accepted for publication 24 February 2014 We re-examined the recent study by Palazzesi et al., (2012) published in the Biological Journal of the Linnean Society (107: 67–85), that presented the historical diversification of Geraniales using BEAST analysis of the plastid spacer trnL–F and of the non-coding nuclear ribosomal internal transcribed spacers (ITS). Their study presented a set of new fossils within the order, generated a chronogram for Geraniales and other rosid orders using fossil-based priors on five nodes, demonstrated an Eocene radiation of Geraniales (and other rosid orders), and argued for more recent (Pliocene–Pleistocene) and climate-linked diversification of genera in the five recognized families relative to previous studies. As a result of very young ages for the crown of Geraniales and other rosid orders, unusual relationships of Geraniales to other rosids, and apparent nucleotide substitution saturation of the two gene regions, we conducted a broad series of BEAST analyses that incorporated additional rosid fossil priors, used more accepted rosid ordinal -

Crocosmia X Crocosmiiflora Montbretia Crocosmia Aurea X Crocosmia Pottsii – Naturally Occurring Hybrid
Top 40 Far Flung Flora A selection of the best plants for pollinators from the Southern Hemisphere List Curated by Thomas McBride From research data collected and collated at the National Botanic Garden of Wales NB: Butterflies and Moths are not studied at the NBGW so any data on nectar plants beneficial for them is taken from Butterfly Conservation The Southern Hemisphere Verbena bonariensis The Southern Hemisphere includes all countries below the equator. As such, those countries are the furthest from the UK and tend to have more exotic and unusual native species. Many of these species cannot be grown in the UK, but in slightly more temperate regions, some species will thrive here and be of great benefit to our native pollinators. One such example is Verbena bonariensis, native to South America, which is a big hit with our native butterfly and bumblebee species. The Southern Hemisphere contains a lower percentage of land than the northern Hemisphere so the areas included are most of South America (particularly Chile, Argentina, Ecuador and Peru), Southern Africa (particularly South Africa) and Oceania (Particularly Australia and New Zealand). A large proportion of the plants in this list are fully hardy in the UK but some are only half-hardy. Half-hardy annuals may be planted out in the spring and will flourish. Half-hardy perennials or shrubs may need to be grown in pots and moved indoors during the winter months or grown in a very sheltered location. The plants are grouped by Tropaeolum majus Continent rather than a full alphabetical -

Updated Angiosperm Family Tree for Analyzing Phylogenetic Diversity and Community Structure
Acta Botanica Brasilica - 31(2): 191-198. April-June 2017. doi: 10.1590/0102-33062016abb0306 Updated angiosperm family tree for analyzing phylogenetic diversity and community structure Markus Gastauer1,2* and João Augusto Alves Meira-Neto2 Received: August 19, 2016 Accepted: March 3, 2017 . ABSTRACT Th e computation of phylogenetic diversity and phylogenetic community structure demands an accurately calibrated, high-resolution phylogeny, which refl ects current knowledge regarding diversifi cation within the group of interest. Herein we present the angiosperm phylogeny R20160415.new, which is based on the topology proposed by the Angiosperm Phylogeny Group IV, a recently released compilation of angiosperm diversifi cation. R20160415.new is calibratable by diff erent sets of recently published estimates of mean node ages. Its application for the computation of phylogenetic diversity and/or phylogenetic community structure is straightforward and ensures the inclusion of up-to-date information in user specifi c applications, as long as users are familiar with the pitfalls of such hand- made supertrees. Keywords: angiosperm diversifi cation, APG IV, community tree calibration, megatrees, phylogenetic topology phylogeny comprising the entire taxonomic group under Introduction study (Gastauer & Meira-Neto 2013). Th e constant increase in knowledge about the phylogenetic The phylogenetic structure of a biological community relationships among taxa (e.g., Cox et al. 2014) requires regular determines whether species that coexist within a given revision of applied phylogenies in order to incorporate novel data community are more closely related than expected by chance, and is essential information for investigating and avoid out-dated information in analyses of phylogenetic community assembly rules (Kembel & Hubbell 2006; diversity and community structure. -

RHS Seed Exchange 2020
RHS Seed Exchange rhs.org.uk/seedlist Introduction to RHS Seed Exchange 2121 The Royal Horticultural Society is the UK’s Dispatch of Orders leading gardening charity, which aims to enrich We will start to send out orders from January everyone’s life through plants, and make the UK a 2020 and dispatch is usually completed by the greener and more beautiful place. This vision end of April. If you have not received your seed underpins all that we do, from inspirational by 1st May please contact us by email: gardens and shows, through our scientific [email protected] research, to our education and community programmes. We’re committed to inspiring Convention on Biological Diversity everyone to grow. 3Nagoya Protocol4 In accordance with the Convention on Biological Most of the seed offered is collected in RHS Diversity (CBD), the Royal Horticultural Society Gardens. Other seed is donated and is offered supplies seed from its garden collections on the under the name provided by the donor. In many conditions that: cases only limited quantities of seed are available. ⅷ The plant material is used for the common However, we feel that even small quantities good in areas of research, education, should be distributed if at all possible. conservation and the development of horticultural institutions or gardens. Our seed is collected from open-pollinated If the recipient seeks to commercialise the plants, therefore may not come true. ⅷ genetic material, its products or resources derived from it, then written permission must Please note we are only able to send seed to be sought from the Royal Horticultural addresses in the UK and EU6 including Society. -

Phylogeny and Historical Biogeography of Geraniaceae In
Systematic Botany (2008), 33(2): pp. 326–342 © Copyright 2008 by the American Society of Plant Taxonomists Phylogeny and Historical Biogeography of Geraniaceae in Relation to Climate Changes and Pollination Ecology Omar Fiz, Pablo Vargas, Marisa Alarcón, Carlos Aedo, José Luis García, and Juan José Aldasoro1 Real Jardín Botanico de Madrid, CSIC, Plaza de Murillo 2, 28014 Madrid, Spain 1Author for correspondence ([email protected]) Communicating Editor: Mark P. Simmons Abstract—Chloroplast (trnL–F and rbcL) sequences were used to reconstruct the phylogeny of Geraniaceae and Hypseocharitaceae. According to these data Hypseocharitaceae and Geraniaceae are monophyletic. Pelargonium and Monsonia are sisters to the largest clade of Geraniaceae, formed by Geranium, Erodium and California. According to molecular dating and dispersal-vicariance analysis, the split of the stem branches of Geraniaceae probably occurred during the Oligocene, in southern Africa or in southern Africa plus the Mediterranean area. However, their diversification occurred during the Miocene, coinciding with the beginning of major aridification events in their distribution areas. An ancestor of the largest clade of Geraniaceae (Geranium, Erodium, and California) colonised a number of habitats in the northern hemisphere and in South American mountain ranges. In summary, the evolution of the Geraniaceae is marked by the dispersal of ancestors from Southern Africa to cold, temperate and often disturbed habitats in the rest of world, where only generalist pollination and facultative autogamy could ensure sufficient seed production and survival. Keywords—autocompatibility, dispersal-vicariance, drought-tolerance, molecular dating, nectaries, P/O indexes. The Geraniaceae are included in the order Geraniales along are characteristic of the Afro-Arabian land mass (Hutchin- with the families Francoaceae, Greyiaceae, Ledocarpaceae, son 1969). -
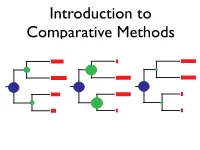
BM CC EB What Can We Learn from a Tree?
Introduction to Comparative Methods BM CC EB What can we learn from a tree? Net diversification (r) Relative extinction (ε) Peridiscaceae Peridiscaceae yllaceae yllaceae h h atop atop Proteaceae Proteaceae r r Ce Ce Tr oc T ho r M M o de c y y H H C C h r r e e a D D a o o nd o e e G G a a m m t t a a d r P r P h h e e u u c c p A p A r e a a e e a a a c a c n n a i B i B h h n d m d l m a l a m m e a e a e e t t n n c u u n n i i d i i e e e e o n o n p n p n a e a S e e S e e n n x x i i r c c a a n o n o p p h g e h g ae e l r a l r a a a a a a i i a a a e a e i b i b h y d c h d c i y i c a a c x c x c c G I a G I a n c n c c c y l y l t a a t a a e e e e e i l c i l c m l m l e c e c f a e a a f a e a a l r r l c c a i i r l e t e t a a r l a a e e u u u u o a o a a a c a c a a l a l e e e b b a a a a e e c e e c a a s c s c c e l c e l e e g e g e a a a a e e n n s e e s e e e e a a a P a P e e N N u u u S u S a e a e a a e e c c l n a l n e e a e e a e a e a e a e r a r a c c C i C i R R a e a e a e a e r c r c A A a d a a d a e i e i phanopetalaceae s r e ph s r e a a s e c s e c e e u u b a a b a e e P P r r l l e n e a a a a m m entho e e e Ha a H o a c r e c r e nt B B e p e e e c e e c a c c h e a p a a p a lo lo l l a a e s o t e s i a r a i a r r r r r a n e a n e a b a l b t a t gaceae e g e ceae a c a s c s a z e z M i a e M i a c a d e a d e ae e ae r e r a e a e a a a c c ce r e r L L i i ac a Vitaceae Vi r r C C e e ta v e v e a a c a a e ea p e ap c a c a e a P P e e l l e Ge G e e ae a t t e e p p r r ce c an u an -

Supplementary Information
Supporting Information for Cornwell et al. 2014. Functional distinctiveness of major 1 plant lineages. doi: 10.1111/1365-2745.12208. SUPPORTING INFORMATION Summary In this Supporting Information, we report additional methods details with respect to the trait data, the climate data, and the analysis. In addition, to more fully explain the method, we include several additional figures. We show the top-five lineages, including the population of lineages from which they were selected, for five important traits (Figure S1 on five separate pages); the bivariate distribution of three selected clades with respect to SLA and leaf N, components of the leaf economic spectrum (Figure S2), the geographic distribution of the clades where this proved useful for interpretation (Figure S3); the procedure for selecting the top six nodes in the leaf N trait, to illustrate the internal behavior of our new comparative method, especially the relative contribution of components of extremeness and the sample size weighting to the results (Figure S4). Finally, we provide references for data used in analyses. SUPPORTING METHODS TRAITS DATABASE We compiled a database for five plant functional traits. Each of these traits is the result of a separate research initiative in which data were gathered directly from researchers leading those efforts and/or the literature; in most, but not all cases, these data have been published elsewhere. Detailed methods for data collection and assembly for each trait are available in the original publications; further data were added for some traits from the primary literature (for references see description of individual traits and Supporting References below). For our compilation, all data were brought to common units for a given trait and thoroughly error checked.