Quantifying the Origins of Life on a Planetary Scale
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Intelligent Design, Abiogenesis, and Learning from History: Dennis R
Author Exchange Intelligent Design, Abiogenesis, and Learning from History: Dennis R. Venema A Reply to Meyer Dennis R. Venema Weizsäcker’s book The World View of Physics is still keeping me very busy. It has again brought home to me quite clearly how wrong it is to use God as a stop-gap for the incompleteness of our knowledge. If in fact the frontiers of knowledge are being pushed back (and that is bound to be the case), then God is being pushed back with them, and is therefore continually in retreat. We are to find God in what we know, not in what we don’t know; God wants us to realize his presence, not in unsolved problems but in those that are solved. Dietrich Bonhoeffer1 am thankful for this opportunity to nature, is the result of intelligence. More- reply to Stephen Meyer’s criticisms over, this assertion is proffered as the I 2 of my review of his book Signature logical basis for inferring design for the in the Cell (hereafter Signature). Meyer’s origin of biological information: if infor- critiques of my review fall into two gen- mation only ever arises from intelli- eral categories. First, he claims I mistook gence, then the mere presence of Signature for an argument against bio- information demonstrates design. A few logical evolution, rendering several of examples from Signature make the point my arguments superfluous. Secondly, easily: Meyer asserts that I have failed to refute … historical scientists can show that his thesis by not providing a “causally a presently acting cause must have adequate alternative explanation” for the been present in the past because the origin of life in that the few relevant cri- proposed candidate is the only known tiques I do provide are “deeply flawed.” cause of the effect in question. -

The Hidden Kingdom
INTRODUCTION Fungi—The Hidden Kingdom OBJECTIVE • To provide students with basic knowledge about fungi Activity 0.1 BACKGROUND INFORMATION The following text provides an introduction to the fungi. It is written with the intention of sparking curiosity about this GRADES fascinating biological kingdom. 4-6 with a K-3 adaptation TEACHER INSTRUCTIONS TYPE OF ACTIVITY 1. With your class, brainstorm everything you know about fungi. Teacher read/comprehension 2. For younger students, hand out the question sheet before you begin the teacher read and have them follow along and MATERIALS answer the questions as you read. • copies of page 11 3. For older students, inform them that they will be given a • pencils brainteaser quiz (that is not for evaluation) after you finish reading the text. VOCABULARY 4. The class can work on the questions with partners or in groups bioremediation and then go over the answers as a class. Discuss any chitin particularly interesting facts and encourage further fungi independent research. habitat hyphae K-3 ADAPTATION kingdom 1. To introduce younger students to fungi, you can make a KWL lichens chart either as a class or individually. A KWL chart is divided moulds into three parts. The first tells what a student KNOWS (K) mushrooms about a subject before it is studied in class. The second part mycelium tells what the student WANTS (W) to know about that subject. mycorrhizas The third part tells what the child LEARNED (L) after studying nematodes that subject. parasitic fungi 2. Share some of the fascinating fungal facts presented in the photosynthesis “Fungi—The Hidden Kingdom” text with your students. -

Framing Major Prebiotic Transitions As Stages of Protocell Development: Three Challenges for Origins-Of-Life Research
Framing major prebiotic transitions as stages of protocell development: three challenges for origins-of-life research Ben Shirt-Ediss1, Sara Murillo-Sánchez2,3 and Kepa Ruiz-Mirazo*2,3 Commentary Open Access Address: Beilstein J. Org. Chem. 2017, 13, 1388–1395. 1Interdisciplinary Computing and Complex BioSystems Group, doi:10.3762/bjoc.13.135 University of Newcastle, UK, 2Dept. Logic and Philosophy of Science, University of the Basque Country, Spain and 3Biofisika Institute Received: 16 February 2017 (CSIC, UPV-EHU), Spain Accepted: 27 June 2017 Published: 13 July 2017 Email: Kepa Ruiz-Mirazo* - [email protected] This article is part of the Thematic Series "From prebiotic chemistry to molecular evolution". * Corresponding author Guest Editor: L. Cronin Keywords: functional integration; origins of life; prebiotic evolution; protocells © 2017 Shirt-Ediss et al.; licensee Beilstein-Institut. License and terms: see end of document. Abstract Conceiving the process of biogenesis as the evolutionary development of highly dynamic and integrated protocell populations provides the most appropriate framework to address the difficult problem of how prebiotic chemistry bridged the gap to full-fledged living organisms on the early Earth. In this contribution we briefly discuss the implications of taking dynamic, functionally inte- grated protocell systems (rather than complex reaction networks in bulk solution, sets of artificially evolvable replicating molecules, or even these same replicating molecules encapsulated in passive compartments) -

Origins of Life in the Universe Zackary Johnson
11/4/2007 Origins of Life in the Universe Zackary Johnson OCN201 Fall 2007 [email protected] Zackary Johnson http://www.soest.hawaii.edu/oceanography/zij/education.html Uniiiversity of Hawaii Department of Oceanography Class Schedule Nov‐2Originsof Life and the Universe Nov‐5 Classification of Life Nov‐7 Primary Production Nov‐9Consumers Nov‐14 Evolution: Processes (Steward) Nov‐16 Evolution: Adaptation() (Steward) Nov‐19 Marine Microbiology Nov‐21 Benthic Communities Nov‐26 Whale Falls (Smith) Nov‐28 The Marine Food Web Nov‐30 Community Ecology Dec‐3 Fisheries Dec‐5Global Ecology Dec‐12 Final Major Concepts TIMETABLE Big Bang! • Life started early, but not at the beginning, of Earth’s Milky Way (and other galaxies formed) history • Abiogenesis is the leading hypothesis to explain the beginning of life on Earth • There are many competing theories as to how this happened • Some of the details have been worked out, but most Formation of Earth have not • Abiogenesis almost certainly occurred in the ocean 20‐15 15‐94.5Today Billions of Years Before Present 1 11/4/2007 Building Blocks TIMETABLE Big Bang! • Universe is mostly hydrogen (H) and helium (He); for Milky Way (and other galaxies formed) example –the sun is 70% H, 28% He and 2% all else! Abundance) e • Most elements of interest to biology (C, N, P, O, etc.) were (Relativ 10 produced via nuclear fusion Formation of Earth Log at very high temperature reactions in large stars after Big Bang 20‐13 13‐94.7Today Atomic Number Billions of Years Before Present ORIGIN OF LIFE ON EARTH Abiogenesis: 3 stages Divine Creation 1. -

The Concept of Organism, a Historical and Conceptual Critique
Do organisms have an ontological status? Charles T. Wolfe Unit for History and Philosophy of Science University of Sydney [email protected] forthcoming in History and Philosophy of the Life Sciences 32:2-3 (2010) Abstract The category of „organism‟ has an ambiguous status: is it scientific or is it philosophical? Or, if one looks at it from within the relatively recent field or sub-field of philosophy of biology, is it a central, or at least legitimate category therein, or should it be dispensed with? In any case, it has long served as a kind of scientific “bolstering” for a philosophical train of argument which seeks to refute the “mechanistic” or “reductionist” trend, which has been perceived as dominant since the 17th century, whether in the case of Stahlian animism, Leibnizian monadology, the neo-vitalism of Hans Driesch, or, lastly, of the “phenomenology of organic life” in the 20th century, with authors such as Kurt Goldstein, Maurice Merleau-Ponty, and Georges Canguilhem. In this paper I try to reconstruct some of the main interpretive „stages‟ or „layers‟ of the concept of organism in order to critically evaluate it. How might „organism‟ be a useful concept if one rules out the excesses of „organismic‟ biology and metaphysics? Varieties of instrumentalism and what I call the „projective‟ concept of organism are appealing, but perhaps ultimately unsatisfying. 1. What is an organism? There have been a variety of answers to this question, not just in the sense of different definitions (an organism is a biological individual; it is a living being, or at least the difference between a living organism and a dead organism is somehow significant in a way that does not seem to make sense for other sorts of entities, like lamps and chairs; it is a self-organizing, metabolic system; etc.) but more tendentiously, in the 1 sense that philosophers, scientists, „natural philosophers‟ and others have both asserted and denied the existence of organisms. -

Bacteria – Friend Or Foe?
Bacteria – Friend or Foe? By Rachel L. Dittmar & Rosa M. Santana Carrero [email protected], [email protected] April 2020 Danger! Sick! Gross! Vaccine! Disease! Medicine! Dirty! MRSA! Eww! E. coli! What is the first thing that comes to mind when you hear “bacteria”? Science! Microscope! MRSA! Salmonella! Hospital! Germs! Staph! Food! Small! 2 Introduction to Bacteria 3 Bacterial nomenclature • Bacteria are referred to by their genus and species, with genus coming first and species coming last: Escherichia coli Escherichia: genus Species: coli Bacteria names are ALWAYS italicized. Genus names are capitalized and species names are not. Sometimes, the genus is abbreviated by its first initial: E. coli 4 Bacteria are prokaryotes. 5 What are prokaryotes? • Plasma membrane separates the cell from its surrounding environment • Cytoplasm contains organelles • Contain DNA consisting of a single large, circular chromosome • Ribosomes make proteins 6 Prokaryotes v. Eukaryotes 7 Image from: https://www.difference.wiki/prokaryotic-cell-vs-eukaryotic-cell/ How do these organisms differ? Prokaryotes Eukaryotes - circular DNA - linear DNA (found in nucleus) - no nucleus - nucleus - no membrane bound organelles - membrane bound organelles - small- less than 10μm - larger than 10μm - unicellular - can be unicellular and multicellular 8 Bacteria are very small. 9 How big are bacteria? Bacteria are very small: 0.1 – 5.0 micrometers. A micrometer (μm) is 0.000001 meters or 0.001 millimeters (mm) For comparison, a human hair is 30 – 100 μm Image from: https://www.khanacademy.org/science/high-school-biology/hs-cells/hs-prokaryotes-and-eukaryotes/a/prokaryotic-cells 10 Bacteria are classified by phenotype or genotype. -

Single-Celled Organisms
Single-Celled Organisms • Humans are made up of millions of tiny cells (multi-celled organisms). • Microorganisms are made up of only ONE cell (single-celled organisms) 1.Single-celled organisms are simple life forms Include: bacteria, protists and others. They move, find food, grow, and make new organisms. 2. Bacteria are one-celled organisms that do NOT have a nucleus. The material that is usually in a nucleus can be found all over the cytoplasm instead. More bacteria on earth than any other living thing Some bacteria are helpful, and some are harmful. One handful of soil can contain billions of bacteria. 3. Protists are a kind of single-celled organism. They live everywhere including pond water, saltwater, animals, and humans Amoebas have a cell membrane, cytoplasm, and a nucleus They move by: changing their shape. 4. Algae are plant-like organisms that Live in water, moist places, and under rocks Among the oldest things on earth! Algae have a cell membrane, cytoplasm, nucleus, and chloroplast. Algae produce most of the oxygen that you breathe! 5. How do single-celled organisms move? Tail-like parts called flagella Hair like structures called cilia Paramecium also use their cilia to get food Multi-Celled Organisms Multi-celled organism is made of up of more than one cell. Plants, animals and other living things. Made up of different kinds of cells that have certain jobs to do. Muscle cells, white blood cells, and nerve cells For example: the cells that make up your heart work together to move oxygen and blood to other cells throughout your body. -

Biology 197: Principles of Organismal Biology Section B: T, R 9:30–10:45 AM Section D: T, R 1:30–2:45 PM Spring 2011 Birck Hall 003 Instructor: Dr
Biology 197: Principles of Organismal Biology Section B: T, R 9:30–10:45 AM Section D: T, R 1:30–2:45 PM Spring 2011 Birck Hall 003 Instructor: Dr. Phil Novack‐Gottshall Office: Birck 332 E‐mail: Blackboard mail preferred Office hours: T 11–1:30, W 11–1, R 11–12:30 (or pnovack‐[email protected]) or by appointment Course Description Organismal biology is one of the major branches of biology and is concerned with all aspects of the life of organisms, including their biodiversity, anatomical structure, physiology, development, biogeography, and ecology. This course is an introductory course required for all biological sciences majors, but it is also useful for gaining basic biological literacy and for those pursuing careers in human and veterinary medicine, psychiatry, agriculture, forestry, microbiology, conservation, ecology, paleontology, environmental science, law, political science, and even cooking, cheese making, and brewing of alcohol. In this class, we will learn the major groups of animals, fungi, plants, protists, algae, and bacteria; their basic characteristics; and how biologists study these organisms to understand their rich evolutionary history, ecological interactions, amazing adaptations, and relevance to humans and other species. In particular, you will practice learning how to view the world and to think like an organismal biologist. Learning objectives 1) Explain the scientific method of organismal biologists to understand the natural world. 2) Identify the major lineages of life through study of their biodiversity, anatomy, physiology, development, behavior, biogeography, fossil record, and ecology. 3) Explain the significance of major transitions in organismal evolution: photosynthesis, endosymbiosis, sexual reproduction, multicellularity, skeletonization, cephalization, terrestrialization, mobility, and carnivory, among others. -

Identification of Substance P Precursor Forms in Human Brain Tissue
Proc. Nati. Acad. Sci. USA Vol. 82, pp. 3921-3924, June 1985 Neurobiology Identification of substance P precursor forms in human brain tissue (prohormones/tachykinins/hypothalamus/substantia nigra/caudate nucleus) FRED NYBERG, PIERRE LE GREvls, AND LARS TERENIUS Department of Pharmacology, University of Uppsala, S-751 24 Uppsala, Sweden Communicated by Tomas Hokfelt, January 28, 1985 ABSTRACT Substance P prohormones were identified in glycine (19). Enzymes involved in the processing of the the caudate nucleus, hypothalamus, and substantia nigra of precursors are incompletely known. human brain. A polypeptide fraction of acidic brain extracts The present paper reports the identification of substance P was fractionated on Sephadex G-50. The Iyophilized fractions precursors in human brain. The strategy for the experimental were sequentially treated with trypsin and a substance P- approach is based on the enzymatic generation in vitro of a degrading enzyme with strong preference toward the Phe7- unique peptide fragment (20, 21). Trypsin treatment of any Phe' and Phe8-Gly9 bonds. The released substance P(1-7) product of the preprotachykinins containing the substance P fragment was isolated by ion-exchange chromatography and sequence generates a fragment with the NH2-terminal region quantitated by a specific radioimmunoassay. Confirmation of identical with that of the native peptide. Further conversion the structure of the isolated radioimmunoassay-active frag- of this fragment with an endopeptidase capable of hydrolyz- ment was achieved by electrophoresis and HPLC. By using this ing substance P at the Phe7-Phe8 bond releases the substance enzymatic/radioimmunoassay procedure, two polypeptide P(1-7) sequence as a fragment that can be recovered by fractions of apparent M, 5000 and 15,000, respectively, were ion-exchange chromatography and quantitated by a specific identified. -
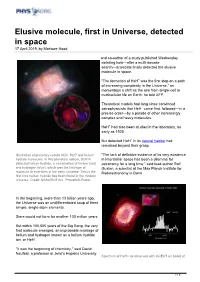
Elusive Molecule, First in Universe, Detected in Space 17 April 2019, by Marlowe Hood
Elusive molecule, first in Universe, detected in space 17 April 2019, by Marlowe Hood and co-author of a study published Wednesday detailing how—after a multi-decade search—scientists finally detected the elusive molecule in space. "The formation of HeH+ was the first step on a path of increasing complexity in the Universe," as momentous a shift as the one from single-cell to multicellular life on Earth, he told AFP. Theoretical models had long since convinced astrophysicists that HeH+ came first, followed—in a precise order—by a parade of other increasingly complex and heavy molecules. HeH+ had also been studied in the laboratory, as early as 1925. But detected HeH+ in its natural habitat had remained beyond their grasp. Illustration of planetary nebula NGC 7027 and helium "The lack of definitive evidence of its very existence hydride molecules. In this planetary nebula, SOFIA in interstellar space has been a dilemma for detected helium hydride, a combination of helium (red) astronomy for a long time," said lead author Rolf and hydrogen (blue), which was the first type of Gusten, a scientist at the Max Planck Institute for molecule to ever form in the early universe. This is the Radioastronomy in Bonn. first time helium hydride has been found in the modern universe. Credit: NASA/SOFIA/L. Proudfit/D.Rutter In the beginning, more than 13 billion years ago, the Universe was an undifferentiated soup of three simple, single-atom elements. Stars would not form for another 100 million years. But within 100,000 years of the Big Bang, the very first molecule emerged, an improbable marriage of helium and hydrogen known as a helium hydride ion, or HeH+. -

The Dissipative Photochemical Origin of Life: UVC Abiogenesis of Adenine
entropy Article The Dissipative Photochemical Origin of Life: UVC Abiogenesis of Adenine Karo Michaelian Department of Nuclear Physics and Applications of Radiation, Instituto de Física, Universidad Nacional Autónoma de México, Circuito Interior de la Investigación Científica, Cuidad Universitaria, Mexico City, C.P. 04510, Mexico; karo@fisica.unam.mx Abstract: The non-equilibrium thermodynamics and the photochemical reaction mechanisms are described which may have been involved in the dissipative structuring, proliferation and complex- ation of the fundamental molecules of life from simpler and more common precursors under the UVC photon flux prevalent at the Earth’s surface at the origin of life. Dissipative structuring of the fundamental molecules is evidenced by their strong and broad wavelength absorption bands in the UVC and rapid radiationless deexcitation. Proliferation arises from the auto- and cross-catalytic nature of the intermediate products. Inherent non-linearity gives rise to numerous stationary states permitting the system to evolve, on amplification of a fluctuation, towards concentration profiles providing generally greater photon dissipation through a thermodynamic selection of dissipative efficacy. An example is given of photochemical dissipative abiogenesis of adenine from the precursor HCN in water solvent within a fatty acid vesicle floating on a hot ocean surface and driven far from equilibrium by the incident UVC light. The kinetic equations for the photochemical reactions with diffusion are resolved under different environmental conditions and the results analyzed within the framework of non-linear Classical Irreversible Thermodynamic theory. Keywords: origin of life; dissipative structuring; prebiotic chemistry; abiogenesis; adenine; organic molecules; non-equilibrium thermodynamics; photochemical reactions Citation: Michaelian, K. The Dissipative Photochemical Origin of MSC: 92-10; 92C05; 92C15; 92C40; 92C45; 80Axx; 82Cxx Life: UVC Abiogenesis of Adenine. -

NASA Armstrong X-Press, May 2019
National Aeronautics and Space Administration Volume 61 Number 4 May 2019 Molecule is found Long search for the universe’s first type of molecule is over By Kassandra Bell and Alison Hawkes USRA and NASA Ames Public Affairs The first type of molecule that ever formed in the universe has been detected in space for the first time, after decades of searching. Scientists discovered its signature in the Milky Way Galaxy using the world’s largest airborne observatory, NASA’s Stratospheric Observatory for Infrared Astronomy, or SOFIA, as the aircraft flew high above the Earth’s surface and pointed its sensitive instruments out into the cosmos. NASA/ESA/Hubble Processing: Judy Schmidt When the universe was still very young, only a few kinds of atoms This image of planetary nebula NGC 7027, with an illustration of helium existed. Scientists believe that around 100,000 years after the big bang, hydride molecules, shows where SOFIA detected helium hydride, a combination helium and hydrogen combined to make a molecule called helium hydride of helium (red) and hydrogen (blue). It was the first type of molecule to ever for the first time. Helium hydride should be present in some parts of the form in the early universe. This is the first time helium hydride has been found modern universe, but it has never been detected in space until now. in the modern universe. SOFIA found modern helium hydride in a planetary nebula, a remnant of what was once a sun-like star. Located 3,000 light-years away near the constellation Cygnus, this planetary nebula, called NGC 7027, has conditions that allow this mystery molecule to form.