Enhanced Arctic-Tethys Connectivity Ended the Toarcian Oceanic Anoxic Event in NW Europe
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
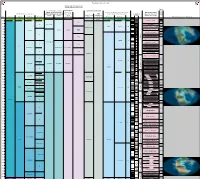
Expanded Jurassic Timescale
TimeScale Creator 2012 chart Russian and Ural regional units Russia Platform regional units Calca Jur-Cret boundary regional Russia Platform East Asian regional units reous stages - British and Boreal Stages (Jur- Australia and New Zealand regional units Marine Macrofossils Nann Standard Chronostratigraphy British regional Boreal regional Cret, Perm- Japan New Zealand Chronostratigraphy Geomagnetic (Mesozoic-Paleozoic) ofossil stages stages Carb & South China (Neogene & Polarity Tethyan Ammonoids s Ma Period Epoch Age/Stage Substage Cambrian) stages Cret) NZ Series NZ Stages Global Reconstructions (R. Blakey) Ryazanian Ryazanian Ryazanian [ no stages M17 CC2 Cretaceous Early Berriasian E Kochian Taitai Um designated ] M18 CC1 145 Berriasella jacobi M19 NJT1 Late M20 7b 146 Lt Portlandian M21 Durangites NJT1 M22 7a 147 Oteke Puaroan Op M22A Micracanthoceras microcanthum NJT1 Penglaizhenian M23 6b 148 Micracanthoceras ponti / Volgian Volgian Middle M24 Burckhardticeras peroni NJT1 Tithonian M24A 6a 149 M24B Semiformiceras fallauxi NJT15 M25 b E 150 M25A Semiformiceras semiforme NJT1 5a Early M26 Semiformiceras darwini 151 lt-Oxf N M-Sequence Hybonoticeras hybonotum 152 Ohauan Ko lt-Oxf R Kimmeridgian Hybonoticeras beckeri 153 m- Lt Late Oxf N Aulacostephanus eudoxus 154 m- NJT14 Late Aspidoceras acanthicum Oxf R Kimmeridgian Kimmeridgian Kimmeridgian Crussoliceras divisum 155 155.431 Card- N Ataxioceras hypselocyclum 156 E Early e-Oxf Sutneria platynota R Idoceras planula Suiningian 157 Cal- Oxf N Epipeltoceras bimammatum 158 lt- Lt Callo -

INFORME July 2014
Chapter 3: Baseline EIA Espejo de Tarapacá Region of Tarapacá, Chile INFORME July 2014 Prepared by: Environmental management Consultants S. A Father Mariano 103 Of. 307 7500499, Providencia, Chile Phone: + 56 2 719 5600 Fax: + 56 2 235 1100 www.gac.cl Capítulo 3: Línea de Base EIA Proyecto Espejo de Tarapacá Index 3. BASELINE ......................................................................................................................................... 3-1 3.1. Introduction ................................................................................................................................. 3-1 3.2. Physical environment ................................................................................................................. 3-2 3.2.1 Atmosphere ........................................................................................................................ 3-2 3.2.1.1 Meteorology .................................................................................................................... 3-2 3.2.1.2 Air quality ........................................................................................................................ 3-5 3.2.1.3 Noise and vibration ......................................................................................................... 3-9 3.2.1.4 Electromagnetic fields .................................................................................................. 3-29 3.2.1.5 Luminosity .................................................................................................................... -

Austroalpine Liassic Ammonites from the Adnet Formation (Northern Calcareous Alps) 163-211 ©Geol
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Jahrbuch der Geologischen Bundesanstalt Jahr/Year: 1993 Band/Volume: 136 Autor(en)/Author(s): Meister Christian, Böhm Florian Artikel/Article: Austroalpine Liassic Ammonites from the Adnet Formation (Northern Calcareous Alps) 163-211 ©Geol. Bundesanstalt, Wien; download unter www.geologie.ac.at Jb. Geol. B.-A. ISSN 0016-7800 Band 136 S.163-211 Wien, Juli 1993 Austroalpine Liassic Ammonites from the Adnet Formation (Northern Calcareous Alps) By CHRISTIAN MEISTER & FLORIAN BÖHM *) With 14 Text-Figures and 9 Plates Oslerreich Salzburg Oberöslerreich Nördliche Kalkalpen Lias Ammoniten Oslerreichische Karle 1. 50.000 Biostratigraphie BI/1l1er94, 95, 96,126 Palaeogeographie Contents Zusammenfassung 163 Abstract. .. 164 Resume 164 1. Introduction 164 2. Geographical and Geological Framework 164 3. Lithological Description and Qualitative/Quantitative Ammonite Distributions 165 3.1. Schmiedwirt Quarry 165 3.2. Breitenberg Quarry 166 3.3. Rotkogel Outcrop 166 3.4. Rötelstein Outcrop 168 4. Systematic Palaeontology 169 PhylioceratinaARKELL 1950 171 LytoceratinaHYATT1889 174 Ammonitina HYATT 1889 175 5. Biostratigraphical Framework 184 5.1. Sinemurian 184 5.1.1. Early Sinemurian 184 5.1.2. Late Sinemurian 184 5.2. Pliensbachian 184 5.2.1. Early Pliensbachian (Carixian) 184 5.2.2. Late Pliensbachian (Domerian) 186 5.3. Toarcian 186 6. Faunal Composition and Palaeogeographical Remarks 189 7. Conclusion 190 Acknowledgements 190 References 208 Oberostalpine Liasammoniten aus der Adnetformation (Nördlichen Kalkalpen) Zusammenfassung Das Oberostalpin spielt eine Schlüsselrolle für das Verständnis der Verteilungsmuster der jurassischen Ammonitenfaunen und für die Fixierung genauer biostratigraphischer Korrelationen zwischen Tethyaler und Euroborealer Faunenprovinz. -

Stratigraphy, 1968
Stratigraphy, 1968 GEOLOGICAL SURVEY BULLETIN 1274 This volume was published as separate chapters A-Q UNITED STATES DEPARTMENT OF THE INTERIOR WALTER J. HICKEL, Secretary GEOLOGICAL SURVEY William T. Pecora, Director CONTENTS [Letters designate the separately published chapters] (A) Changes in stratigraphic nomenclature by the U.S. Geological Survey, 1967, by George V. Cohee, Robert G. Bates, and Wilna B. Wright. (B) Stratigraphy of the Albemarle Group of the Carolina slate belt in central North Carolina, by Arvid A. Stromquist and Harold W. Sundelius. (C) New and revised stratigraphic names in the Santa Rita Mountains of south- . eastern Arizona, by Harald Drewes. (D) The coal-bearing group in the Nenana coal field, Alaska, by Clyde Wahr- haftig, Jack A. Wolfe, Estella B. Leopold, and Marvin A. Lanphere. (E) Definition of Wisconsinan Stage, by John C. Frye, H. B. Willman, Meyer Rubin, and Robert F. Black. (F) The Reany Creek Formation, Marquette County, Michigan, by Willard P. Puffett. (G) The Aguada Limestone of northwestern Puerto Rico, by Watson H. Monroe. (H) Cretaceous Tertiary boundary in New Jersey, Delaware, and eastern Maryland, by James P. Minard, James P. Owens, Norman F. Sohl, Harold E. Gill, and James F. Mello. (I) Six new Paleozoic and Mesozoic formations in east-central Alaska, by Earl E. Brabb. (J) Four new members of the Upper Cretaceous Straight Cliffs Formation in the southeastern Kaiparowits region, Kane County, Utah, by Fred Peterson. (K) Summary of Cretaceous stratigraphy in part of the McCarthy quadrangle, Alaska., by D. L. Jones and E. M. MacKevett, Jr. (L) The Quimby and Greenvale Cove Formations in western Maine, by Robert H. -

Les Ammonites Domérien Des Causses
Christian MEISTER LES AMMONITES DU DOMÉRIEN DES CAUSSES (FRANCE) cahiers de paléontologie « cahiers de paléontologie • cahiers de paléontologie Éditions du CNRS CAHIERS DE PALÉONTOLOGIE LES AMMONITES DU DOMERIEN DES CAUSSES (FRANCE) Analyses paléontologiques et stratigraphiques par Christian Meister U A 157, Dijon ÉDITIONS DU CENTRE NATIONAL DE LA RECHERCHE SCIENTIFIQUE 15, quai Anatole France — 75700 PARIS 1989 Christian M eister Institut des Sciences de la Terre de l’Université de Bourgogne et Centre de Géodynamique sédimentaire et Evolution géobiologi que, U.A. C.N.R.S. 157, 6 Bd Gabriel 21100 Dijon, dans le cadre d’un travail subventionné par le Fonds National Suisse de la Recherche Scientifique. © Centre National de la Recherche Scientifique, Paris, 1989 ISSN 0766-0502 — ISBN 2-222-4310-7 TABLE DES MATIERES RESUMÉ/ABSTRACT ..................................................................................................................................... 9 I. INTRODUCTION..................................................................................................................................... 11 II. CADRE GÉOGRAPHIQUE ET GÉOLOGIQUE ............................................................................. 12 III. SUCCESSION DES FAUNES D’AMMONITES DOMÉRIENNES ............................................. 14 Le Samonta ............................................................................................................................................... 15 Rivière-sur-Tarn........................................................................................................................................ -

Review of Fossil Collections in Scotland Scotland South Scotland South
Detail of Permian footprints Chelichnus duncani in sandstone from Dumfries and Galloway. Dumfries Museum and Camera Obscura. © Dumfries and Galloway Council Museums Service Review of Fossil Collections in Scotland Scotland South Scotland South Dumfries Museum and Camera Obscura (Dumfries and Galloway Council) Sanquhar Tolbooth Museum (Dumfries and Galloway Council) Stranraer Museum (Dumfries and Galloway Council) Gem Rock Museum Newton Stewart Museum Tweeddale Museum (Live Borders) Hawick Museum (Live Borders) 1 Dumfries Museum and Camera Obscura (Dumfries and Galloway Council) Collection type: Local authority Accreditation: 2018 The Observatory, Rotchell Road, Dumfries, DG2 7SW Contact: [email protected] Location of collections The original Museum was located in a windmill dating from the late 1700s, preserved in the 1830s by the newly-formed Dumfries and Maxwelltown Astronomical Society for use as an observatory. An extension built in the 1860s added a large gallery and mezzanine level with further gallery and storage space added in the 1980s. Collections are onsite in displays and a main storeroom. Size of collections 1,000-1,200 fossils. Onsite records Information is in an Adlib CMS database transcribed from several previous electronic systems and various paper documents, such as MDA and other index card systems, Gift Books, Accession Registers, free text catalogues, inventories and listings. Fossils are catalogued with other geological material as a series of numbered boxes with a list compiled in the 1980s by James Williams (1944- 2009). An online catalogue is available at: https://dgc-web.adlibhosting.com/home but does not yet contain fossil entries. Collection highlights 1. Permian vertebrate trackways from Locharbriggs and Corncockle quarries linked to Reverend Henry Duncan (1774-1846) and former curator Alfred Truckell (1919-2007). -
GEOLOGICAL SURVEY PROFESSIONAL PAPER 1148 Early Jurassic Ammonites from Alaska
GEOLOGICAL SURVEY PROFESSIONAL PAPER 1148 Early Jurassic Ammonites from Alaska By RALPH W. IMLAY GEOLOGICAL SURVEY PROFESSIONAL PAPER 1148 Studies of Early Jurassic ammonites from Alaska provide close correlations with Lower Jurassic ammonite zones in Europe and with ammonite successions elsewhere in the world UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1981 UNITED STATES DEPARTMENT OF THE INTERIOR JAMES G. WATT, Secretary GEOLOGICAL SURVEY Doyle G. Frederick, Acting Director Library of Congress Cataloging in Publication Data Imlay, Ralph Willard, 1908- Early Jurassic ammonites from Alaska. (Geological Survey professional paper ; 1148) Bibliography: p. Supt. of Docs, no.: I 19.16:1148 Includes index. 1. Ammonoidea. 2. Paleontology Jurassic. 3. Paleontology Alaska. I. Title. II. Series. QE807.A51585 564'.53 81-607901 AACR2 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 CONTENTS Page Abstract _____________________________________________——— 1 Introduction ______________________________________________ 2 Biologic analysis ___________________________________________ 2 Biostratigraphic summary _____________________________________ 4 Northern Alaska _________________________________________ 4 East-central Alaska ________________________________________ 6 Southern Alaska _________________________________——————— 7 Wrangell Mountains _____________________________________ 7 Talkeetna Mountains ____________________________________ 7 Chulitna River area _____________________________________ -
Download The
LOWER TO MIDDLE JURASSIC (PLIENSBACHIAN TO BAJOC1AN) STRATIGRAPHY AND PLIENSBACHIAN AMMONITE FAUNA OF THE NORTHERN SPATSIZI AREA, NORTH CENTRAL BRITISH COLUMBIA by ROBERT CHARLES THOMSON B.Sc, UNIVERSITY OF ALBERTA, 1981 A THESIS SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE in THE FACULTY OF GRADUATE STUDIES DEPARTMENT OF GEOLOGICAL SCIENCES We accept this thesis as conforming to the required standard THE UNIVERSITY OF BRITISH COLUMBIA OCTOBER, 1985 ® ROBERT CHARLES THOMSON, 1985 In presenting this thesis in partial fulfilment of the requirements for an advanced degree at the The University of British Columbia, I agree that the Library shall make it freely available for reference and study. I further agree that permission for extensive copying of this thesis for scholarly purposes may be granted by the Head of my Department or by his or her representatives. It is understood that copying or publication of this thesis for financial gain shall not be allowed without my written permission. DEPARTMENT OF GEOLOGICAL SCIENCES The University of British Columbia 2075 Wesbrook Place Vancouver, Canada V6T 1W5 Date: OCTOBER. 1985 Abstract The lithostratigraphy and Pliensbachian ammonite fauna of a sequence of Pliensbachian to Bajocian sedimentary rocks, informally referred to here as the Spatsizi Group, from the Spatsizi map-area (104 H) in north-central British Columbia are examined in this thesis. Twenty Five species of ammonites representing fifteen genera from Pliensbachian rocks of the Spatsizi Group are described and their stratigraphic ranges in the thesis area determined. The Spatsizi fauna is comprised primarily of ammonites of Tethyan aspect and also contains elements endemic to the East Pacific faunal realm. -
Review of Fossil Collections in Scotland Review of Fossil Collections in Scotland
Detail of the Upper Devonian fishHoloptychius from Dura Den, Fife. © Perth Museum & Art Gallery, Perth & Kinross Council Review of Fossil Collections in Scotland Review of Fossil Collections in Scotland Contents Introduction 3 Background 3 Aims of the Collections Review 4 Methodology 4 Terminology 5 Summary of fossil material 6 Influences on collections 14 Collections by region Aberdeen and North East 17 Elgin Museum (Moray Society) 18 Falconer Museum (Moray Council) 21 Stonehaven Tolbooth Museum 23 The Discovery Centre (Live Life Aberdeenshire) 24 Arbuthnot Museum (Live Life Aberdeenshire) 27 Zoology Museum (University of Aberdeen Museums) 28 Meston Science Building (University of Aberdeen Museums) 30 Blairs Museum 37 Highlands and Islands 38 Inverness Museum and Art Gallery (High Life Highland) 39 Nairn Museum 42 West Highland Museum (West Highland Museum Trust) 44 Brora Heritage Centre (Brora Heritage Trust) 45 Dunrobin Castle Museum 46 Timespan (Timespan Heritage and Arts Society) 48 Stromness Museum (Orkney Natural History Society) 50 Orkney Fossil and Heritage Centre 53 Shetland Museum and Archives (Shetland Amenity Trust) 56 Bute Museum (Bute Museum Trust) 58 Hugh Miller’s Birthplace Cottage and Museum (National Trust for Scotland) 59 Treasures of the Earth 62 Staffin Dinosaur Museum 63 Gairloch Museum (Gairloch & District Heritage Company Ltd) 65 Tayside, Central and Fife 66 Stirling Smith Art Gallery and Museum 67 Perth Museum and Art Gallery (Culture Perth and Kinross) 69 The McManus: Dundee’s Art Gallery and Museum (Leisure -

Prehistoric Reptile One Mile Above Mccarthy, Alaska in Wrangell-St
National Park Service U.S. Department of the Interior Natural Resource Stewardship and Science Paleontological Resource Inventory and Monitoring Central Alaska Network Natural Resource Technical Report NPS/NRSS/NRTR—2011/510 Wrangell-Saint Elias National Park and Preserve Wrangell-Saint Elias National Park and Preserve (WRST) was originally proclaimed as a national monument on December 1, 1978. Two years later, on December 2, 1980, the monument was established as a national park and preserve, and a large portion of this expansive park was designated as wilderness. Wrangell-Saint Elias National Park and Preserve represents the largest area of the National Park System and encompasses 5,332,106 ha (13,175,901 acres), larger than nine states and more than twice the land area of Maryland. The park represents 3,368,263 ha (8,323,148 acres) and the preserve represents 1,963,842 ha (4,852,753 acres). Three mountain ranges, the Chugach, Saint Elias, and Wrangell, converge within the park to form what is commonly referred to as the “mountain kingdom of North America.” The park includes the continent’s largest assemblage of glaciers and the greatest collection of peaks above 4,570 m (15,000 ft) in elevation. Mount Saint Elias is the second highest peak in the United States standing at 5,488.8 m (18,008 ft) above sea level. Wrangell-Saint Elias National Park and Preserve was also designated a World Heritage Site on October 24, 1979. WRST is surrounded by other federal and provincial areas both in Alaska and Canada. To the south the park is Glacier Bay National Park and Preserve (see the Southeast Alaska Network Paleontological Resource Inventory Report; Santucci and Kenworthy 2008). -

INTEGRATED AMMONITE BIOCEIRONOLOGY and U-Pb GEOCHRONOLOGY from a BASAL JURASSIC SECTION in ALASKA
National Liirary Bibliothèque nationale I*I of Canada du Canada Acquisitions and Acquisitions et Bibliographie Services services bibliographiques 395 Wellington Street 395. rue Wellington OltawaûN KlAON4 Ollawa ON K1A ON4 Canada Canada The author has granted a non- L'auteur a accordé une licence non exclusive licence allowing the exclusive permettant à la National Library of Canada to Bi1,liothèque nationale du Canada de reproduce, loan, distribute or sell reproduire, prêter, distribuer ou copies of this thesis in microform, vendre des copies de cette thése sous paper or electronic formats. la forme de microfiche/film, de reproduction sur papier ou sur format électronique. The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts fiom it Ni la thèse ni des extraits substantiels may be printed or othexwise de ceile-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation. Current tirne scales disagree on age estirnates for Jurassic stage boundaries and carry large uncertainties. The U-Pb ~r"~~r-"~rdating of voIcaniclastic rocks of preciseiy known biochronologic age is the preferred method to improve the calibtation. Eighteen new U-Pb zircon dates were obtained on volcaniclastic rock from Surassic volcanic arc assemblages of the North American Cordillera. The voIcanic rocks are interbedded in fossiliferous marine sedimenq rocks, which in mm were dated by ammonite biochronology at the zona1 level. In the Queen Charlotte Islands, a tuff Iayer directly beneath the Triassic-Jurassic boundary was dated at 200I1 Ma. -

Preliminary Bedrock Geology of the Mcbride River Area (Parts of NTS
British Columbia Geological Survey 129°10' Open File 2018-3 Grid north COL ISH UM Turnagain Lake IT B R IA B Magnetic Preliminary bedrock geology of True north 490,000 mE north G the McBride River area E Y O E 6,460,000 mN V L R O GICAL SU PzJCC 129°05' Location map Magnetic lJTs declination Parts of NTS Sheets 104H/14,15; 104I/02, 03, 06 19° 11.40' E Previous mapping in the region YT Convergence Atlin angle YT Terranes (after Nelson et al., 2013) Bram I. van Straaten and Sebastian J. Bichlmaier 0° 4.26' W This study CC Ancestral North America 58.5 ° 104J/10 104J/09 104I/12 104I/11 104I/10 104I/09 Intermontane terranes Dease Lake Approximate mean declination 2018 van Straaten et al. (2017) Stikinia (ST), Cache Creek (CC), Quesnellia for centre of map Dease Lake (QN), Yukon-Tanana (YT) and others Annual change, decreasing 17.2' Logan et al. (2012) Recommended citation Coast plutonic complex van Straaten, B.I. and Bichlmaier, S.J., 2018. Preliminary bedrock geology of the McBride River area. British Columbia Ministry of Energy, Mines and Petroleum Resources, British Columbia Geological Survey Open File 2018-3, 1:50,000 scale. ST Insular terranes van Straaten et al. (2012) 104J/07 104J/08 104I/05 104I/06 104I/07 104I/08 58°15' Stewart Outboard terranes 58°15' Evenchick and Thorkelson (2005) mJHMUvm 500,000 mE Geochronology data 0 2 4 km 1400 37 Gabrielse (1998) QN Sample Easting Northing Unit Age (Ma) Type Mineral Sample description Source Smithers 104J/02 104J/01 104I/02 104I/01 58 ° 104I/04 104I/03 Read and Psutka (1990) AN-78-467 493616 6434342 JMRgd 166±8 U-Pb Zrn Hbl-Bt-bearing granodiorite Anderson et al.