Association Between ABO and Duffy Blood Types and Circulating Chemokines and Cytokines
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Hemolytic Disease of the Newborn
Intensive Care Nursery House Staff Manual Hemolytic Disease of the Newborn INTRODUCTION and DEFINITION: Hemolytic Disease of the Newborn (HDN), also known as erythroblastosis fetalis, isoimmunization, or blood group incompatibility, occurs when fetal red blood cells (RBCs), which possess an antigen that the mother lacks, cross the placenta into the maternal circulation, where they stimulate antibody production. The antibodies return to the fetal circulation and result in RBC destruction. DIFFERENTIAL DIAGNOSIS of hemolytic anemia in a newborn infant: -Isoimmunization -RBC enzyme disorders (e.g., G6PD, pyruvate kinase deficiency) -Hemoglobin synthesis disorders (e.g., alpha-thalassemias) -RBC membrane abnormalities (e.g., hereditary spherocytosis, elliptocytosis) -Hemangiomas (Kasabach Merritt syndrome) -Acquired conditions, such as sepsis, infections with TORCH or Parvovirus B19 (anemia due to RBC aplasia) and hemolysis secondary to drugs. ISOIMMUNIZATION A. Rh disease (Rh = Rhesus factor) (1) Genetics: Rh positive (+) denotes presence of D antigen. The number of antigenic sites on RBCs varies with genotype. Prevalence of genotype varies with the population. Rh negative (d/d) individuals comprise 15% of Caucasians, 5.5% of African Americans, and <1% of Asians. A sensitized Rh negative mother produces anti-Rh IgG antibodies that cross the placenta. Risk factors for antibody production include 2nd (or later) pregnancies*, maternal toxemia, paternal zygosity (D/D rather than D/d), feto-maternal compatibility in ABO system and antigen load. (2) Clinical presentation of HDN varies from mild jaundice and anemia to hydrops fetalis (with ascites, pleural and pericardial effusions). Because the placenta clears bilirubin, the chief risk to the fetus is anemia. Extramedullary hematopoiesis (due to anemia) results in hepatosplenomegaly. -

Blood Product Replacement: Obstetric Hemorrhage
CMQCC OBSTETRIC HEMORRHAGE TOOLKIT Version 2.0 3/24/15 BLOOD PRODUCT REPLACEMENT: OBSTETRIC HEMORRHAGE Richard Lee, MD, Los Angeles County and University of Southern California Medical Center Laurence Shields, MD, Marian Regional Medical Center/Dignity Health Holli Mason, MD, Cedars-Sinai Medical Center Mark Rollins, MD, PhD, University of California, San Francisco Jed Gorlin, MD, Innovative Blood Resources/Memorial Blood Center, St. Paul, Minnesota Maurice Druzin, MD, Lucile Packard Children’s Hospital Stanford University Jennifer McNulty, MD, Long Beach Memorial Medical Center EXECUTIVE SUMMARY • Outcomes are improved with early and aggressive intervention. • Both emergency blood release and massive transfusion protocols should be in place. • In the setting of significant obstetric hemorrhage, resuscitation transfusion should be based on vital signs and blood loss and should not be delayed by waiting for laboratory results. • Calcium replacement will often be necessary with massive transfusion due to the citrate used for anticoagulation in blood products. • During massive transfusion resuscitation, the patient’s arterial blood gas, electrolytes, and core temperature should be monitored to guide clinical management and all transfused fluids should be warmed; direct warming of the patient should be initiated as needed to maintain euthermia and to avoid added coagulopathy. BACKGROUND AND LITERATURE REVIEW After the first several units of packed red blood cells (PRBCs) and in the face of continuing or worsening hemorrhage, aggressive transfusion therapy becomes critical. This report covers the experience with massive transfusion protocols. Lessons from military trauma units as well as civilian experience with motor vehicle accidents and massive obstetric hemorrhage have identified new principles such as earlier use of plasma (FFP/thawed plasma/plasma frozen within 24 hours/liquid plasma) and resuscitation transfusion while laboratory results are pending. -
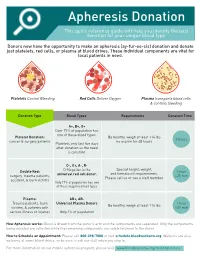
Apheresis Donation This Quick Reference Guide Will Help You Identify the Best Donation for Your Unique Blood Type
Apheresis Donation This quick reference guide will help you identify the best donation for your unique blood type. Donors now have the opportunity to make an apheresis (ay-fur-ee-sis) donation and donate just platelets, red cells, or plasma at blood drives. These individual components are vital for local patients in need. Platelets Control Bleeding Red Cells Deliver Oxygen Plasma transports blood cells & controls bleeding Donation Type Blood Types Requirements Donation Time A+, B+, O+ Over 75% of population has one of these blood types. Platelet Donation: Be healthy, weigh at least 114 lbs 2 hours cancer & surgery patients no aspirin for 48 hours Platelets only last five days after donation so the need is constant. O-, O+, A-, B- Special height, weight, Double Red: O-Negative is the 1 hour and hematocrit requirements. surgery, trauma patients, universal red cell donor. +25 min Please call us or see a staff member accident, & burn victims Only 17% of population has one of these negative blood types Plasma: AB+, AB- Trauma patients, burn Universal Plasma Donors 1 hour Be healthy, weigh at least 114 lbs victims, & patients with +30 min serious illness or injuries Only 4% of population How Apheresis works: Blood is drawn from the donor’s arm and the components are separated. Only the components being donated are collected while the remaining components are safely returned to the donor How to Schedule an Appointment: Please call 800-398-7888 or visit schedule.bloodworksnw.org. Walk-ins are also welcome at some blood drives, so be sure to ask our staff when you stop in. -

Guidelines for Transfusion and Patient Blood Management, and Discuss Relevant Transfusion Related Topics
Guidelines for Transfusion and Community Transfusion Committee Patient Blood Management Community Transfusion Committee CHAIR: Aina Silenieks, M.D., [email protected] MEMBERS: A.Owusu-Ansah, M.D. S. Dunder, M.D. M. Furasek, M.D. D. Lester, M.D. D. Voigt, M.D. B. J. Wilson, M.D. COMMUNITY Juliana Cordero, Blood Bank Coordinator, CHI Health Nebraska Heart REPRESENTATIVES: Becky Croner, Laboratory Services Manager, CHI Health St. Elizabeth Mackenzie Gasper, Trauma Performance Improvement, Bryan Medical Center Kelly Gillaspie, Account Executive, Nebraska Community Blood Bank Mel Hanlon, Laboratory Specialist - Transfusion Medicine, Bryan Medical Center Kyle Kapple, Laboratory Quality Manager, Bryan Medical Center Lauren Kroeker, Nurse Manager, Bryan Medical Center Christina Nickel, Clinical Laboratory Director, Bryan Medical Center Rachael Saniuk, Anesthesia and Perfusion Manager, Bryan Medical Center Julie Smith, Perioperative & Anesthesia Services Director, Bryan Medical Center Elaine Thiel, Clinical Quality Improvement/Trans. Safety Officer, Bryan Med Center Kelley Thiemann, Blood Bank Lead Technologist, CHI Health St. Elizabeth Cheryl Warholoski, Director, Nebraska Operations, Nebraska Community Blood Bank Jackie Wright, Trauma Program Manager, Bryan Medical Center CONSULTANTS: Jed Gorlin, M.D., Innovative Blood Resources [email protected] Michael Kafka, M.D., LifeServe Blood Center [email protected] Alex Smith, D.O., LifeServe Blood Center [email protected] Nancy Van Buren, M.D., Innovative -

Blood Type and Transplantation & A2 Donor to B Recipient
Page 1 of 2 Blood Type and Transplantation Information for Kidney Transplant Patients Does blood type matter in transplantation? Everyone waiting for a transplant has their blood typed. You will have one of four blood types: O, A, B or AB. Your blood type is determined by the antigens that are present on your blood cells. These antigens are A or B. These antigens will be found both in your blood and on your organs. What antigen does each blood type have? Blood type O Blood type A Blood type B Blood type AB have no have A antigens. have B antigens. have both A O antigens. A B AB and B antigens. How does my body react to antigens? Your body will react to antigens that are different than your own by attacking with antibodies. Antibodies are proteins created by your immune system to attack anything that does not belong. Antibodies are the soldiers in your body’s army protecting you from foreign invasions such as viruses. Unfortunately, the antibodies cannot tell the difference between harmful viruses and beneficial transplanted organs. What blood type will my donor be? Transplants can occur between all blood types. However, when the donor’s blood type is different than yours and there are different antigens being transplanted on your new organ, your antibodies will be triggered and attack the transplanted organ. This is called rejection. Because of this, transplants usually happen between a donor and a recipient of the same blood type. This is called an identical transplant. Can I get an organ from a donor that has a different blood type than mine? Yes! If you do not have antibodies in your body against the antigens that come from the donor, your immune system should not attack the transplanted organ. -

The Johns Hopkins Comprehensive Transplant Center Incompatible Kidney Transplant Programs
The Johns Hopkins Comprehensive Transplant Center Incompatible Kidney Transplant Programs The Johns Hopkins Comprehensive Transplant Center’s Incompatible Kidney Transplantation Program allows many patients previously thought to be “incompatible” to receive the gift of life. The program is comprised of several elements: Blood Type Incompatible Kidney Transplant Program Positive Crossmatch and Sensitized Patient Program Paired Kidney Exhange Program Altruistic Donor Program Blood Type Incompatible Kidney Transplant Program More than one-third of willing live donors are turned down because their blood types are not compatible with the person to whom they wish to donate their kidney. Most of us have natural antibodies against organs from people with different blood types. These antibodies can rapidly destroy a transplanted kidney. The Blood Type Incompatible Transplant Program allows patients to receive a kidney from a live donor who has an incompatible blood type (see fig1.). Patients in this program must be willing to undergo all prescribed treatments before and after the transplant to remove harmful antibodies and decrease the risk of rejection. Figure 1: Blood Type Compatibility Chart Donor Recipient How are harmful antibodies removed? Harmful antibodies are removed with a process called plasmapheresis, a procedure similar to dialysis that removes the plasma portion of the blood where antibodies are located. The number of plasmapheresis treatments required by the recipient before surgery varies depending on the amount of harmful antibodies in their blood. After each plasmapheresis the recipient receives an intravenous infusion of immune globulin to replace antibodies needed to fight infections and help prevent harmful antibodies from returning. Once the antibodies against the donor’s blood type decrease to very low levels, the transplantation can take place. -

Therapeutic Apheresis, J Clin Apheresis 2007, 22, 104-105
Apheresis: Basic Principles, Practical Considerations and Clinical Applications Joseph Schwartz, MD Anand Padmanabhan, MD PhD Director, Transfusion Medicine Assoc Med Director/Asst Prof Columbia Univ. Medical Center BloodCenter of Wisconsin New York Presbyterian Hospital Medical College of Wisconsin Review Session, ASFA Annual meeting, Scottsdale, Arizona, June 2011 Objectives (Part 1) • Mechanism of Action • Definitions • Technology (ies) • Use • Practical Considerations • Math • Clinical applications – HPC Collection Objectives (Part 2) • Clinical applications: System/ Disease Specific Indications • ASFA Fact Sheet Apheresis •Derives from Greek, “to carry away” •A technique in which whole blood is taken and separated extracorporealy, separating the portion desired from the remaining blood. •This allows the desired portion (e.g., plasma) to be removed and the reminder returned. Apheresis- Mechanism of Action •Large-bore intravenous catheter connected to a spinning centrifuge bowl •Whole blood is drawn from donor/patient into the centrifuge bowl •The more dense elements, namely the RBC, settle to the bottom with less dense elements such as WBC and platelets overlying the RBC layer and finally, plasma at the very top. Apheresis: Principles of Separation Platelets (1040) Lymphocytes Torloni MD (1050-1061) Monocytes (1065 - 1069) Granulocyte (1087 - 1092) RBC Torloni MD Torloni MD Separate blood components is based on density with removal of the desired component Graphics owned by and courtesy of Gambro BCT Principals of Apheresis WBC Plasma Torlo RBC ni MD Torloni MD RBC WBC Plasma G Cobe Spectra Apheresis- Mechanism of Action Definitions • Plasmapheresis: plasma is separated, removed (i.e. less than 15% of total plasma volume) without the use of replacement solution • Plasma exchange (TPE): plasma is separated, removed and replaced with a replacement solution such as colloid (e.g. -

Direct Antibody Test (DAT) Positive
Direct antibody test (DAT) positive Information for patients, parents and guardians As part of your routine pregnancy screening, you However, only a small number of DAT positive have had a direct antibody test (DAT). We’ve babies will develop these problems. Babies given you this factsheet to explain what a DAT who are not DAT positive can still develop involves and what it means if your baby is DAT anaemia and jaundice. A positive DAT simply positive. If you have any further questions, tells us to look out for any signs of anaemia and please speak to a member of your healthcare jaundice. It does not necessarily mean that your team who will be pleased to advise you. baby will need treatment. What is a direct antibody test (DAT)? If we find out that you have rhesus negative During pregnancy, some of the blood between (Rh-) blood during pregnancy, we may give the mother and baby may mix. This mixing of you an injection called anti-D to stop your blood sometimes produces antibodies (proteins body making antibodies against your baby’s that are part of your body’s natural defences) blood. Occasionally this injection causes the which may become a problem for the baby. DAT result to come out positive. Babies who are DAT positive for this reason do not usually As part of routine antenatal (pregnancy) develop anaemia or jaundice. screening blood tests, your midwife will record your blood type (blood group) and check What is anaemia? whether your blood contains any antibodies Anaemia is a very common condition where the that may affect your baby’s red blood cells. -

Guide to Blood Plasma Products and Their Applications – Part One
Vet Times The website for the veterinary profession https://www.vettimes.co.uk Guide to blood plasma products and their applications – part one Author : Amanda Boag, Jenny Walton Categories : Vets Date : September 26, 2011 Amanda Boag and Jenny Walton discuss indications for the use of plasma products, their production, and storage within the practice in the first of a three-part series FOLLOWING changes in legislation in 2005, veterinary blood banking and component therapy has developed rapidly within the UK. Practitioners now have easy access to canine blood and its products – notably, packed red cells and various plasma products. This series of articles will focus on indications for plasma product use, their production and storage, the administration and monitoring of plasma transfusions and will include a series of five case studies that have used plasma products. Indications for plasma component therapy Although well-defined indications exist for the use of plasma products in single or multiple coagulation deficiencies, indications for many of its other uses are empiric. Haemostatic disorders Plasma components are most commonly used to treat haemostatic disorders and are generally effective in treating both inherited and acquired conditions. Specific diagnosis enables selection of the optimum replacement product. It is important to remember that any samples drawn for diagnostic purposes (for example, measurement of clotting factor levels) must be taken either 1 / 5 before or at least 36 hours after transfusion to allow the measurement of endogenous levels. Inherited bleeding disorders Plasma components are administered to control active haemorrhage or as preoperative prophylaxis. Inherited bleeding disorders are associated with deficiency of a specific factor and the optimal product for treatment depends on which factor is lacking. -

Blood Collection and Handling Tube Additives, Tube Type Most Laboratory Tests Are Performed on Plasma, Serum, Or Whole Blood
Blood Collection and Handling Tube Additives, Tube Type Most laboratory tests are performed on plasma, serum, or whole blood. To preserve the specimen in the form required by the test, collection tubes contain additives that either prevent coagulation (for plasma and whole blood recovery), or activate coagulation (for serum recovery). Please refer to individual test requirements. Drawing Order When multiple tubes are drawn, it is important to prioritize the drawing order to prevent a Table 1. Vacutainer Order Of Draw tube additive from contaminating the next tube and altering the chemical composition of the 1. Navy Blue (metals testing) following specimen. Coagulation tests are highly susceptible to interference from 2. Blood culture bottles / SPS tubes contamination from tissue fluid and tube additives; therefore these tests are usually collected 3. Coagulation tests: first when a series of tubes are collected. Prior to collecting tests for coagulation (i.e. Blue top a. Clear top “waste” tube tube) a plain Clear top tube containing no additive must be partially filled and discarded. b. Light Blue top This “waste” tube prevents tissue thromboplastins from contaminating the Blue top tube. Blue top tubes must be allowed to fill to the line indicated on the tube, exhausting the 4. Gold top vacuum. See Table 1, “Vacutainer Order of Draw” for proper collection order of vacutainer 5. Plain Red top tubes. 6. Dark Green top (heparin) 7. Light Green top Certain blood collection techniques have been identified as possible sources of error in 8. Lavender or Pink top laboratory testing. Avoid the following sources of test error when collecting blood: 9. -

A Fatal Case of Severe Hemolytic Disease of Newborn Associated with Anti-Jkb
J Korean Med Sci 2006; 21: 151-4 Copyright � The Korean Academy ISSN 1011-8934 of Medical Sciences A Fatal Case of Severe Hemolytic Disease of Newborn Associated with Anti-Jkb The Kidd blood group is clinically significant since the Jk antibodies can cause acute Won Duck Kim, Young Hwan Lee* and delayed transfusion reactions as well as hemolytic disease of newborn (HDN). In general, HDN due to anti-Jkb incompatibility is rare and it usually displays mild Department of Pediatrics, Dongguk University, College of Medicine, Gyeongju; Department of Pediatrics*, clinical symptoms with a favorable prognosis. Yet, we apparently experienced the Yeungnam University, College of Medicine, Daegu, second case of HDN due to anti-Jkb with severe clinical symptoms and a fatal out- Korea come. A female patient having the AB, Rh(D)-positive boodtype was admitted for jaundice on the fourth day after birth. At the time of admission, the patient was lethar- gic and exhibited high pitched crying. The laboratory data indicated a hemoglobin value of 11.4 mg/dL, a reticulocyte count of 14.9% and a total bilirubin of 46.1 mg/dL, Received : 18 October 2004 a direct bilirubin of 1.1 mg/dL and a strong positive result (+++) on the direct Coomb’s Accepted : 11 February 2005 test. As a result of the identification of irregular antibody from the maternal serum, anti-Jkb was detected, which was also found in the eluate made from infant’s blood. Despite the aggressive treatment with exchange transfusion and intensive photother- apy, the patient died of intractable seizure and acute renal failure on the fourth day Address for correspondence of admission. -

Neonatal Unconjugated Hyperbilirubinemia: Risk Assessment and Management
Neonatal Unconjugated Hyperbilirubinemia: Risk Assessment and Management SHEENA BHAVSAR, PA-C PEDIATRIC FACULTY BAYLOR COLLEGE OF MEDICINE Objectives . Importance and Impact on infant health . Physiologic and Pathologic causes of hyperbilirubinemia in newborns . Guide to a systematic risk assessment . Management . Follow up . Common parent/provider questions Unconjugated Hyperbilirubinemia . Elevated levels of bilirubin in the blood, >95th percentile based to the hour specific nomogram . Severe hyperbilirubinemia is defined as > 25mg/dL . One of the primary causes of hospital readmission of neonates . Usually peaks between 3-5 days of life Bilirubin Encephalopathy - highly neurotoxic substance - can bind to brain tissue and cause neurologic dysfunction . ACUTE (hypotonia, lethargy, poor suck irritability, high pitched cry, intermittent hypertonia , seizures, fever, apnea, coma) VS . CHRONIC (movement disorders, auditory neuropathy, oculomotor dysfunction, GI concerns) KERNICTERUS Causes? INCREASED ERYTHROCYTE DECREASED BILIRUBIN BREAKDOWN CLEARANCE . Physiologic Jaundice . Physiologic Jaundice . Trauma . Breastmilk Jaundice . Blood Incompatibility . Anatomic Obstruction . Infection . Liver enzyme defect . Enzyme Deficiency . Drug Induced . Globin Synthesis Defect . Liver Disease . Membrane conditions . Metabolic INCREASED ERYTHROCYTE DECREASED BILIRUBIN BREAKDOWN CLEARANCE . Physiologic Jaundice . Physiologic Jaundice . Trauma . Breastmilk Jaundice . Blood Incompatibility . Anatomic Obstruction . Infection . Liver enzyme defect . Enzyme