PRA on Apriona Species
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

11-16589 PRA Record Saperda Candida
PRA Saperda candida 11-16589 (10-15760, 09-15659) European and Mediterranean Plant Protection Organisation Organisation Européenne et Méditerranéenne pour la Protection des Plantes Guidelines on Pest Risk Analysis Decision-support scheme for quarantine pests Version N°3 Pest Risk Analysis for Saperda candida Pest risk analysts: Expert Working group for PRA for Saperda candida (met in 2009-11) ANDERSON Helen (Ms) - The Food and Environment Research Agency (GB) AGNELLO Arthur (Mr) - Department of Entomology New York State Agricultural Experiment Station(USA) BAUFELD Peter (Mr) - Julius Kühn Institut (JKI), Federal Research Centre for Cultivated Plants, Institute for National and International Plant Health (DE) GILL Bruce D. (Mr) - Head Entomology, Ottawa Plant Laboratories, C.F.I.A. (CA) PFEILSTETTER Ernst (Mr) - Julius Kühn Institut (JKI), Federal Research Centre for Cultivated Plants, Institute for National and International Plant Health (DE) (core member) STEFFEK Robert (Mr) - Austrian Agency for Health and Food Safety (AGES), Institute for Plant Health (AT) (core member) VAN DER GAAG Dirk Jan (Mr) - Plant Protection Service (NL) (core member) The Section on risk management was reviewed by the EPPO Panel on Phytosanitary Measures in 2010-02-18. 1 PRA Saperda candida Stage 1: Initiation 1 Identification of a In summer 2008, the presence of Saperda candida was detected for the first time in Germany and in Europe (Nolte & Give the reason for performing the PRA single pest Krieger, 2008). This wood boring insect was observed on the island of Fehmarn on urban trees ( Sorbus intermedia and other host plants) and eradication measures were taken against it. S. candida is considered as a pest of apple trees and other tree species in North America. -

31 First Record of Batocera Rufomaculata (De Geer, 1775) from Sunderban Biosphere Reserve, West Bengal
International Journal of Entomology Research ISSN: 2455-4758 www.entomologyjournals.com Volume 1; Issue 3; March 2016; Page No. 31-32 First record of Batocera rufomaculata (De Geer, 1775) from Sunderban biosphere reserve, West Bengal 1 Bulganin Mitra, 2 Udipta Chakraborti, 3 Olive Biswas, 4 Sankarsan Roy, 5 Kaushik Mallick, 6 Priyanka Das 1, 2, 3, 4, 6 Zoological Survey of India, Prani Vigyan Bhawan, M-Block, New Alipore, Kolkata. 5 Post Graduate Department of Zoology, Asutosh College, Kolkata Abstract Studies on Longhorn beetles (Coleoptera) in Sunderban region is very poor. Altogether, 8 species under 3 subfamilies are already reported from Sunderban Biosphere Reserve. Present communication reports Batocera rufomaculata (De Geer, 1775) for the first time from this Biosphere reserve. Keywords: Sunderban Biosphere Reserve, Cerambycidae, Lamiinae, Batocera Introduction Sunderban region in India is 9600 sq km (4200 sq km of Reserved Forest and 5400 sq km of non-forest, inhabited region) which constitutes the Sunderban Biosphere Reserve (SBR). Indian Sunderban is bound on the west by river Muriganga and on the east by rivers Harinbhahga and Raimangal. Administrative boundary of the Sunderban is spread over two districts i.e. North 24-Parganas (Hingalganj, Hasnabad, Haroa, Sandeskhali - I,II, and Minakhan blocks) and South 24-Parganas (Sagar, Namkhana, Kakdwip, Patharpratima, Kultali, Mathurapur-I,II, Jaynagar-I,II, Canning-I,II, Basanti and Gosaba blocks).The extent of mangrove Reserve Forests in Indian Sunderban is around 4260 sq km, out of which 55% is under land vegetation cover and balance 45% is under water body/ inter-tidal zone. Studies on beetles and weevils (Coleoptera) in Sunderban region is very poor. -

4 Reproductive Biology of Cerambycids
4 Reproductive Biology of Cerambycids Lawrence M. Hanks University of Illinois at Urbana-Champaign Urbana, Illinois Qiao Wang Massey University Palmerston North, New Zealand CONTENTS 4.1 Introduction .................................................................................................................................. 133 4.2 Phenology of Adults ..................................................................................................................... 134 4.3 Diet of Adults ............................................................................................................................... 138 4.4 Location of Host Plants and Mates .............................................................................................. 138 4.5 Recognition of Mates ................................................................................................................... 140 4.6 Copulation .................................................................................................................................... 141 4.7 Larval Host Plants, Oviposition Behavior, and Larval Development .......................................... 142 4.8 Mating Strategy ............................................................................................................................ 144 4.9 Conclusion .................................................................................................................................... 148 Acknowledgments ................................................................................................................................. -

Transcriptome and Gene Expression Analysis of Rhynchophorus Ferrugineus (Coleoptera: Curculionidae) During Developmental Stages
Transcriptome and gene expression analysis of Rhynchophorus ferrugineus (Coleoptera: Curculionidae) during developmental stages Hongjun Yang1,2, Danping Xu1, Zhihang Zhuo1,2,3, Jiameng Hu2 and Baoqian Lu4 1 College of Life Science, China West Normal University, Nanchong, Sichuan, China 2 Key Laboratory of Genetics and Germplasm Innovation of Tropical Special Forest Trees and Ornamental Plants, Ministry of Education, Key Laboratory of Germplasm Resources Biology of Tropical Special Ornamental Plants of Hainan Province, College of Forestry, Hainan University, Haikou, Hainan, China 3 Key Laboratory of Integrated Pest Management on Crops in South China, Ministry of Agriculture, South China Agricultural University, Guangzhou, Guangdong, China 4 Key Laboratory of Integrated Pest Management on Tropical Crops, Ministry of Agriculture China, Environ- ment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan, China ABSTRACT Background. Red palm weevil, Rhynchophorus ferrugineus Olivier, is one of the most destructive pests harming palm trees. However, genomic resources for R. ferrugineus are still lacking, limiting the ability to discover molecular and genetic means of pest control. Methods. In this study, PacBio Iso-Seq and Illumina RNA-seq were used to generate transcriptome from three developmental stages of R. ferrugineus (pupa, 7th-instar larva, adult) to increase the understanding of the life cycle and molecular characteristics of the pest. Results. Sequencing generated 625,983,256 clean reads, from which 63,801 full-length transcripts were assembled with N50 of 3,547 bp. Expression analyses revealed 8,583 differentially expressed genes (DEGs). Moreover, gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis revealed that these Submitted 5 March 2020 Accepted 29 September 2020 DEGs were mainly related to the peroxisome pathway which associated with metabolic Published 2 November 2020 pathways, material transportation and organ tissue formation. -

Proceedings, 23Rd U.S. Department of Agriculture Interagency Research
United States Department of Proceedings Agriculture 23rd U.S. Department of Agriculture Forest Service Northern Interagency Research Forum on Research Station Invasive Species 2012 General Technical Report NRS-P-114 The findings and conclusions of each article in this publication are those of the individual author(s) and do not necessarily represent the views of the U.S. Department of Agriculture or the Forest Service. All articles were received in digital format and were edited for uniform type and style. Each author is responsible for the accuracy and content of his or her paper. The use of trade, firm, or corporation names in this publication is for the information and convenience of the reader. Such use does not constitute an official endorsement or approval by the U.S. Department of Agriculture or the Forest Service of any product or service to the exclusion of others that may be suitable. This publication reports research involving pesticides. It does not contain recommendations for their use, nor does it imply that the uses discussed here have been registered. All uses of pesticides must be registered by appropriate State and/or Federal, agencies before they can be recommended. CAUTION: Pesticides can be injurious to humans, domestic animals, desirable plants, and fi sh or other wildlife—if they are not handled or applied properly. Use all pesticides selectively and carefully. Follow recommended practices for the disposal of surplus pesticides and pesticide containers. Cover graphic by Vincent D’Amico, U.S. Forest Service, Northern Research Station. Manuscript received for publication August 2012 Published by: For additional copies: U.S. -

Arthropods of Elm Fork Preserve
Arthropods of Elm Fork Preserve Arthropods are characterized by having jointed limbs and exoskeletons. They include a diverse assortment of creatures: Insects, spiders, crustaceans (crayfish, crabs, pill bugs), centipedes and millipedes among others. Column Headings Scientific Name: The phenomenal diversity of arthropods, creates numerous difficulties in the determination of species. Positive identification is often achieved only by specialists using obscure monographs to ‘key out’ a species by examining microscopic differences in anatomy. For our purposes in this survey of the fauna, classification at a lower level of resolution still yields valuable information. For instance, knowing that ant lions belong to the Family, Myrmeleontidae, allows us to quickly look them up on the Internet and be confident we are not being fooled by a common name that may also apply to some other, unrelated something. With the Family name firmly in hand, we may explore the natural history of ant lions without needing to know exactly which species we are viewing. In some instances identification is only readily available at an even higher ranking such as Class. Millipedes are in the Class Diplopoda. There are many Orders (O) of millipedes and they are not easily differentiated so this entry is best left at the rank of Class. A great deal of taxonomic reorganization has been occurring lately with advances in DNA analysis pointing out underlying connections and differences that were previously unrealized. For this reason, all other rankings aside from Family, Genus and Species have been omitted from the interior of the tables since many of these ranks are in a state of flux. -

Oregon Invasive Species Action Plan
Oregon Invasive Species Action Plan June 2005 Martin Nugent, Chair Wildlife Diversity Coordinator Oregon Department of Fish & Wildlife PO Box 59 Portland, OR 97207 (503) 872-5260 x5346 FAX: (503) 872-5269 [email protected] Kev Alexanian Dan Hilburn Sam Chan Bill Reynolds Suzanne Cudd Eric Schwamberger Risa Demasi Mark Systma Chris Guntermann Mandy Tu Randy Henry 7/15/05 Table of Contents Chapter 1........................................................................................................................3 Introduction ..................................................................................................................................... 3 What’s Going On?........................................................................................................................................ 3 Oregon Examples......................................................................................................................................... 5 Goal............................................................................................................................................................... 6 Invasive Species Council................................................................................................................. 6 Statute ........................................................................................................................................................... 6 Functions ..................................................................................................................................................... -

25Th U.S. Department of Agriculture Interagency Research Forum On
US Department of Agriculture Forest FHTET- 2014-01 Service December 2014 On the cover Vincent D’Amico for providing the cover artwork, “…and uphill both ways” CAUTION: PESTICIDES Pesticide Precautionary Statement This publication reports research involving pesticides. It does not contain recommendations for their use, nor does it imply that the uses discussed here have been registered. All uses of pesticides must be registered by appropriate State and/or Federal agencies before they can be recommended. CAUTION: Pesticides can be injurious to humans, domestic animals, desirable plants, and fish or other wildlife--if they are not handled or applied properly. Use all pesticides selectively and carefully. Follow recommended practices for the disposal of surplus pesticides and pesticide containers. Product Disclaimer Reference herein to any specific commercial products, processes, or service by trade name, trademark, manufacturer, or otherwise does not constitute or imply its endorsement, recom- mendation, or favoring by the United States government. The views and opinions of wuthors expressed herein do not necessarily reflect those of the United States government, and shall not be used for advertising or product endorsement purposes. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, sex, religion, age, disability, political beliefs, sexual orientation, or marital or family status. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at 202-720-2600 (voice and TDD). To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, Room 326-W, Whitten Building, 1400 Independence Avenue, SW, Washington, D.C. -

Catalogue of Afghanistan Longhorn Beetles (Coleoptera, Cerambycidae) with Two Descriptions of New Phytoecia (Parobereina Danilevsky, 2018) from Central Asia
Humanity space International almanac VOL. 8, No 2, 2019: 104-140 http://zoobank.org/urn:lsid:zoobank.org:pub:30F6FA0A-2D7A-4ED2-9EAE-AB7707FFBE61 Catalogue of Afghanistan Longhorn beetles (Coleoptera, Cerambycidae) with two descriptions of new Phytoecia (Parobereina Danilevsky, 2018) from Central Asia M.A. Lazarev State Budget Professional Educational Institution of the Moscow Region “Chekhov technical college” Novaya str., 4, Novyi Byt village, Chekhov District, Moscow Region 142322 Russia e-mail: [email protected]; [email protected] Key words: Coleoptera, Cerambycidae, taxonomy, distribution, new species, Afghanistan, Pakistan. Abstract: The Catalogue includes all 78 Cerambycidae species of Afghanistan fauna known up to 2019 with the references to the original descriptions; 22 species were not mentioned for Afghanistan in Palaearctic Cerambycidae Catalogue by Löbl & Smetana (2010). Bibliography of each species usually includes the geographical information from corresponding publications. Many new taxonomy positions published after 2010 are used here without special remarks. Agapanthia (Epoptes) dahli ustinovi Danilevsky, 2013 stat. nov. is downgraded from the species level. Two species are described as new Phytoecia (Parobereina) pashtunica sp. n. from Afghanistan and Phytoecia (Parobereina) heinzi sp.n. from Pakistan. The present work is an attempt to summarize all data published up to now on Cerambycidae of Afghanistan fauna. Family CERAMBYCIDAE Latreille, 1802 subfamily Prioninae Latreille, 1802 tribe Macrotomini J. Thomson, 1861 genus Anomophysis Quentin & Villiers, 1981: 374 type species Prionus spinosus Fabricius, 1787 inscripta C.O. Waterhouse, 1884: 380 (Macrotoma) Heyrovský, 1936: 211 - Wama; Tippmann, 1958: 41 - Kabul, Ost- Afghanistan, 1740; Sarobi, am Kabulflus, 900 m; Mangul, Bashgultal, Nuristan, Ost-Afghanistan, 1250 m; Fuchs, 1961: 259 - Sarobi 1100 m, O.-Afghanistan; Fuchs, 1967: 432 - Afghanistan, 25 km N von Barikot, 1800 m, Nuristan; Nimla, 40 km SW von Dschelalabad; Heyrovský, 1967: 156 - Zentral-Afghanistan, Prov. -
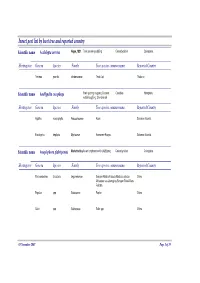
Insect Pest List by Host Tree and Reported Country
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae -

Scrub Oak Preserve Animal Checklist Volusia County, Florida
Scrub Oak Preserve Animal Checklist Volusia County, Florida Accipitridae Cervidae Cooper's Hawk Accipiter cooperii White-tailed Deer Odocoileus virginianus Red-tailed Hawk Buteo jamaicensis Red-shouldered Hawk Buteo lineatus Charadriidae Bald Eagle Haliaeetus leucocephalus Killdeer Charadrius vociferus Acrididae Corvidae American Bird Grasshopper Schistocerca americana Scrub Jay Aphelocoma coerulescens American Crow Corvus brachyrhynchos Agelenidae Fish Crow Corvus ossifragus Grass Spider Agelenopsis sp. Blue Jay Cyanocitta cristata Anatidae Dactyloidae Wood Duck Aix sponsa Cuban Brown Anole Anolis sagrei Blattidae Elateridae Florida woods cockroach Eurycotis floridana Eyed Click Beetle Alaus oculatus Bombycillidae Emberizidae Cedar Waxwing Bombycilla cedrorum Eastern Towhee Pipilo erythrophthalmus Cardinalidae Eumenidae Northern Cardinal Cardinalis cardinalis Paper wasp Polistes sp. Cathartidae Formicidae Turkey Vulture Cathartes aura Carpenter ants Camponotus sp. Black Vulture Coragyps atratus Fire Ant Solenopsis invicta Certhiidae Fringillidae Blue-gray Gnatcatcher Polioptila caerulea American Goldfinch Carduelis tristis Geomyidae Phalacrocoracidae Southeastern Pocket Gopher Geomys pinetis Double-crested Cormorant Phalacrocorax auritus Gruidae Picidae Sandhill Crane Grus canadensis Northern Flicker Colaptes auratus Pileated Woodpecker Dryocopus pileatus Hesperiidae Red-bellied Woodpecker Melanerpes carolinus Duskywing Erynnis sp. Downy Woodpecker Picoides pubescens Hirundinidae Polychrotidae Tree Swallow Tachycineta bicolor Green Anole -

EPPO Reporting Service
ORGANISATION EUROPEENNE EUROPEAN AND ET MEDITERRANEENNE MEDITERRANEAN POUR LA PROTECTION DES PLANTES PLANT PROTECTION ORGANIZATION EPPO Reporting Service NO. 4 PARIS, 2018-04 General 2018/068 New data on quarantine pests and pests of the EPPO Alert List 2018/069 Quarantine lists of Kazakhstan (2017) 2018/070 EPPO report on notifications of non-compliance 2018/071 EPPO communication kits: templates for pest-specific posters and leaflets 2018/072 Useful publications on Spodoptera frugiperda Pests 2018/073 First report of Tuta absoluta in Tajikistan 2018/074 First report of Tuta absoluta in Lesotho 2018/075 First reports of Grapholita packardi and G. prunivora in Mexico 2018/076 First report of Scaphoideus titanus in Ukraine 2018/077 First report of Epitrix hirtipennis in France 2018/078 First report of Lema bilineata in Italy 2018/079 Eradication of Anoplophora glabripennis in Brünisried, Switzerland 2018/080 Update on the situation of Anoplophora glabripennis in Austria Diseases 2018/081 First report of Ceratocystis platani in Turkey 2018/082 Huanglongbing and citrus canker are absent from Egypt 2018/083 Xylella fastidiosa eradicated from Switzerland 2018/084 Update on the situation of Ralstonia solanacearum on roses in Switzerland 2018/085 First report of ‘Candidatus Phytoplasma fragariae’ in Slovenia Invasive plants 2018/086 Ambrosia artemisiifolia control in agricultural areas in North-west Italy 2018/087 Optimising physiochemical control of invasive Japanese knotweed 2018/088 Update on LIFE project IAP-RISK 2018/089 Conference: Management and sharing of invasive alien species data to support knowledge-based decision making at regional level (2018-09-26/28, Bucharest, Romania) 21 Bld Richard Lenoir Tel: 33 1 45 20 77 94 E-mail: [email protected] 75011 Paris Fax: 33 1 70 76 65 47 Web: www.eppo.int EPPO Reporting Service 2018 no.