In Vitro Kinetic Characterization of Axitinib Metabolism S
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Effects of Combination Treatments on Drug Resistance in Chronic Myeloid Leukaemia: an Evaluation of the Tyrosine Kinase Inhibitors Axitinib and Asciminib H
Lindström and Friedman BMC Cancer (2020) 20:397 https://doi.org/10.1186/s12885-020-06782-9 RESEARCH ARTICLE Open Access The effects of combination treatments on drug resistance in chronic myeloid leukaemia: an evaluation of the tyrosine kinase inhibitors axitinib and asciminib H. Jonathan G. Lindström and Ran Friedman* Abstract Background: Chronic myeloid leukaemia is in principle a treatable malignancy but drug resistance is lowering survival. Recent drug discoveries have opened up new options for drug combinations, which is a concept used in other areas for preventing drug resistance. Two of these are (I) Axitinib, which inhibits the T315I mutation of BCR-ABL1, a main source of drug resistance, and (II) Asciminib, which has been developed as an allosteric BCR-ABL1 inhibitor, targeting an entirely different binding site, and as such does not compete for binding with other drugs. These drugs offer new treatment options. Methods: We measured the proliferation of KCL-22 cells exposed to imatinib–dasatinib, imatinib–asciminib and dasatinib–asciminib combinations and calculated combination index graphs for each case. Moreover, using the median–effect equation we calculated how much axitinib can reduce the growth advantage of T315I mutant clones in combination with available drugs. In addition, we calculated how much the total drug burden could be reduced by combinations using asciminib and other drugs, and evaluated which mutations such combinations might be sensitive to. Results: Asciminib had synergistic interactions with imatinib or dasatinib in KCL-22 cells at high degrees of inhibition. Interestingly, some antagonism between asciminib and the other drugs was present at lower degrees on inhibition. -
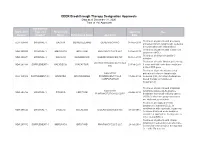
CDER Breakthrough Therapy Designation Approvals Data As of December 31, 2020 Total of 190 Approvals
CDER Breakthrough Therapy Designation Approvals Data as of December 31, 2020 Total of 190 Approvals Submission Application Type and Proprietary Approval Use Number Number Name Established Name Applicant Date Treatment of patients with previously BLA 125486 ORIGINAL-1 GAZYVA OBINUTUZUMAB GENENTECH INC 01-Nov-2013 untreated chronic lymphocytic leukemia in combination with chlorambucil Treatment of patients with mantle cell NDA 205552 ORIGINAL-1 IMBRUVICA IBRUTINIB PHARMACYCLICS LLC 13-Nov-2013 lymphoma (MCL) Treatment of chronic hepatitis C NDA 204671 ORIGINAL-1 SOVALDI SOFOSBUVIR GILEAD SCIENCES INC 06-Dec-2013 infection Treatment of cystic fibrosis patients age VERTEX PHARMACEUTICALS NDA 203188 SUPPLEMENT-4 KALYDECO IVACAFTOR 21-Feb-2014 6 years and older who have mutations INC in the CFTR gene Treatment of previously untreated NOVARTIS patients with chronic lymphocytic BLA 125326 SUPPLEMENT-60 ARZERRA OFATUMUMAB PHARMACEUTICALS 17-Apr-2014 leukemia (CLL) for whom fludarabine- CORPORATION based therapy is considered inappropriate Treatment of patients with anaplastic NOVARTIS lymphoma kinase (ALK)-positive NDA 205755 ORIGINAL-1 ZYKADIA CERITINIB 29-Apr-2014 PHARMACEUTICALS CORP metastatic non-small cell lung cancer (NSCLC) who have progressed on or are intolerant to crizotinib Treatment of relapsed chronic lymphocytic leukemia (CLL), in combination with rituximab, in patients NDA 206545 ORIGINAL-1 ZYDELIG IDELALISIB GILEAD SCIENCES INC 23-Jul-2014 for whom rituximab alone would be considered appropriate therapy due to other co-morbidities -

Nonclinical Antiangiogenesis And
Cancer Therapy: Preclinical Nonclinical Antiangiogenesis and Antitumor Activities of Axitinib (AG-013736), an Oral, Potent, and Selective Inhibitor of Vascular Endothelial Growth Factor Receptor Tyrosine Kinases 1, 2, 3 Dana D. Hu-Lowe, Helen Y.Zou, Maren L. Grazzini, Max E. Hallin, Grant R. Wickman, Karin Amundson, JeffreyH.Chen,DavidA.Rewolinski,ShinjiYamazaki,EllenY.Wu,MicheleA.McTigue,BrionW.Murray, Robert S. Kania, Patrick O’Connor, David R. Shalinsky, and Steve L. Bender Abstract Purpose: Axitinib (AG-013736) is a potent and selective inhibitor of vascular endothelial growth factor (VEGF) receptor tyrosine kinases 1to 3 that is in clinical development for the treatment of solid tumors. We provide a comprehensive description of its in vitro characteristics and activities, in vivo antiangiogenesis, and antitumor efficacy and translational pharmacology data. Experimental Design: The potency, kinase selectivity, pharmacologic activity, and antitumor efficacy of axitinib were assessed in various nonclinical models. Results: Axitinib inhibitscellular autophosphorylation of VEGF receptors(VEGFR) with picomo- lar IC50 values. Counterscreening across multiple kinase and protein panels shows it is selective for VEGFRs. Axitinib blocks VEGF-mediated endothelial cell survival, tube formation, and down- stream signaling through endothelial nitric oxide synthase, Akt and extracellular signal-regulated kinase. Following twice daily oral administration, axitinib produces consistent and dose-depen- dent antitumor efficacy that is associated with blocking VEGFR-2 phosphorylation, vascular per- meability, angiogenesis, and concomitant induction of tumor cell apoptosis. Axitinib in combination with chemotherapeutic or targeted agentsenhancesantitumor efficacy in many tu- mor models compared with single agent alone. Dose scheduling studies in a human pancreatic tumor xenograft model show that simultaneous administration of axitinib and gemcitabine with- out prolonged dose interruption or truncation of axitinib produces the greatest antitumor efficacy. -

Study Protocol);
AG-013736, PF-02341066 A4061068 Final Protocol Amendment 2, 27 January 2016 CLINICAL PROTOCOL A PHASE 1B, OPEN LABEL, DOSE ESCALATION STUDY TO EVALUATE SAFETY, PHARMACOKINETICS AND PHARMACODYNAMICS OF AXITINIB (AG-013736) IN COMBINATION WITH CRIZOTINIB (PF-02341066) IN PATIENTS WITH ADVANCED SOLID TUMORS Compounds: AG-013736, PF-02341066 Compound Names: axitinib, crizotinib United States (US) Investigational New IND 63,662 Drug (IND) Numbers: IND 73,544 European Clinical Trials Database 2015-001724-31 (EudraCT) Number Protocol Number: A4061068 Phase: 1b Page 1 AG-013736, PF-02341066 A4061068 Final Protocol Amendment 2, 27 January 2016 Document History Document Version Date Summary of Changes and Rationale Amendment 2 26 January 2016 Added EudraCT number. SCHEDULE OF ACTIVITIES: - Contraception check added to align with the Sponsor’s most recent protocol template. - Registration and Study Treatment: updated to clarify which patient subgroup will receive axitinib single agent during a 7-day Lead-In period and which subgroup will start with the combination without the Lead-In period. - Archival Tumor Tissue, De Novo Tumor Biopsy: footnotes updated to align with Patient Selection Section, Inclusion Criterion #1. Section 1. Introduction: literature updated; axitinib and crizotinib safety data updated according to the last version of the Investigator’s Brochure. Section 4. Patient Selection: - Inclusion Criterion #1, Dose Expansion Phase: language updated: 1. to make the selected patient population more in line with the actual 2nd and 3rd line advanced RCC population; 2. to include alternative option for the collection of biopsy in the Dose Expansion Cohort 2 patients. - Exclusion Criterion #8: language updated to align with the current standard language of crizotinib clinical protocols. -

Combination Strategy Targeting VEGF and HGF/C-Met in Human Renal
Published OnlineFirst November 7, 2014; DOI: 10.1158/1535-7163.MCT-14-0094 Small Molecule Therapeutics Molecular Cancer Therapeutics Combination Strategy Targeting VEGF and HGF/c-met in Human Renal Cell Carcinoma Models Eric Ciamporcero1,2, Kiersten Marie Miles1, Remi Adelaiye1,3, Swathi Ramakrishnan1,3, Li Shen1, ShengYu Ku1,3, Stefania Pizzimenti2, Barbara Sennino4, Giuseppina Barrera2, and Roberto Pili1 Abstract Alternative pathways to the VEGF, such as hepatocyte growth 1Â/day) of RP-R-01–bearing mice. Treatment with single-agent factor or HGF/c-met, are emerging as key players in tumor crizotinib reduced tumor vascularization but failed to inhibit angiogenesis and resistance to anti-VEGF therapies. The aim of tumor growth in either model, despite also a significant increase this study was to assess the effects of a combination strategy of c-met expression and phosphorylation in the sunitinib-resis- targeting the VEGF and c-met pathways in clear cell renal cell tant tumors. In contrast, axitinib treatment was effective in inhi- carcinoma (ccRCC) models. Male SCID mice (8/group) were biting angiogenesis and tumor growth in both models, with its implanted with 786-O tumor pieces and treated with either a antitumor effect significantly increased by the combined treat- selective VEGF receptor tyrosine kinase inhibitor, axitinib (36 mg/ ment with crizotinib, independently from c-met expression. kg, 2Â/day); a c-met inhibitor, crizotinib (25 mg/kg, 1Â/day); or Combination treatment also induced prolonged survival and combination. We further tested this drug combination in a significant tumor growth inhibition in the 786-O human RCC human ccRCC patient–derived xenograft, RP-R-01, in both model. -

For Health Professionals Who Care for Cancer Patients
May 2014 Volume 17, Number 5 For Health Professionals Who Care For Cancer Patients Inside This Issue: . Editor’s Choice – New Programs: Breast – Trastuzumab Patient Handouts (Chinese, Punjabi): Emtansine for Metastatic Breast Cancer (UBRAVKAD); Cyclophosphamide IV , Etoposide IV, Hydroxyurea Leukemia/BMT – Arsenic Trioxide for Untreated . Benefit Drug List – New: UBRAVKAD, HLHETCSPA, (ULKATOATRA, ULKATOP) and Relapsed (ULKATOR) Acute ULKATOATRA, ULKTOP, ULKTOR, SAAVTC; Revised: Promyelocytic Leukemia; Lymphoma – Etoposide, LYALEM Dexamethasone and Cyclosporine for Hemophagocytic . Systemic Therapy Update – Editorial Board Lymphohistiocytosis (HLHETCSPA); Sarcoma – Topotecan Membership and Cyclophosphamide for Relapsed Neuroblastoma, Ewing’s Sarcoma, Osteogenic Sarcoma or . List of New and Revised Protocols, Provincial Pre‐ Rhabdomyosarcoma (SAAVTC); Highlights of Changes in Printed Orders and Patient Handouts – New: Protocols, PPPOs, and Patient Handouts: Lung – Expanding UBRAVKAD, HLHETCSPA, HNAVP, HNNAVP, Eligibility Criteria of Maintenance Pemetrexed in Advanced UHNNAVPC, ULKATOATRA, ULKTOP, ULKTOR, Non‐Small Cell Lung Cancer (ULUAVPMTN) ULUAVCRIZ, SAAVTC, SCDMAB; Revised: UBRAVERIB, BRAVEXE, BRAVT7, UBRAVLCAP, UBRAVTCAP, . Provincial Systemic Therapy Program – Updates to BCCA CNAJTZRT, CNOCTLAR, GIEFUPRT, GIPAJGEM, GIRCRT, Policy on Physician Coverage for Medical Emergencies GOOVDDCAT, UGUAXIT, UGUPENZ, UGUSORAF, During Delivery of Selective Chemotherapy Drugs HNAVP, UHNLADCF, HNNAVPE, UHNLACETRT, . Communities Oncology Network – -

5.01.534 Multiple Receptor Tyrosine Kinase Inhibitors
PHARMACY POLICY – 5.01.534 Multiple Receptor Tyrosine Kinase Inhibitors Effective Date: Aug. 1, 2021 RELATED MEDICAL POLICIES: Last Revised: July 9, 2021 5.01.517 Use of Vascular Endothelial Growth Factor Receptor (VEGF) Inhibitors and Replaces: N/A Other Angiogenesis Inhibitors in Oncology Patients 5.01.518 BCR-ABL Kinase Inhibitors 5.01.544 Prostate Cancer Targeted Therapies 5.01.589 BRAF and MEK Inhibitors 5.01.603 Epidermal Growth Factor Receptor (EGFR) Inhibitors Select a hyperlink below to be directed to that section. POLICY CRITERIA | CODING | RELATED INFORMATION EVIDENCE REVIEW | REFERENCES | HISTORY ∞ Clicking this icon returns you to the hyperlinks menu above. Introduction An enzyme is a chemical messenger. Tyrosine kinases are enzymes within cells. They serve as on/off switches for many of the cells’ functions. One of their most important roles is to help send signals telling a cell to grow. If there is a genetic change that leaves the switch permanently on, cells grow without stopping and tumors form. Multiple tyrosine kinase inhibitors block the “grow” signal in specific types of tumors. This policy discusses when multiple receptor tyrosine kinase inhibitors may be considered medically necessary. Note: The Introduction section is for your general knowledge and is not to be taken as policy coverage criteria. The rest of the policy uses specific words and concepts familiar to medical professionals. It is intended for providers. A provider can be a person, such as a doctor, nurse, psychologist, or dentist. A provider also can be a place where medical care is given, like a hospital, clinic, or lab. -

Neoadjuvant Combination of Pazopanib Or Axitinib And
2102 Original Article Neoadjuvant combination of pazopanib or axitinib and programmed cell death protein-1-activated dendritic cell-cytokine-induced killer cells immunotherapy may facilitate surgery in patients with renal cell carcinoma Zhiling Zhang1,2,3#, Longbin Xiong1,2,3#, Zeshen Wu1,2,3#, Huiming Liu2,3,4, Kang Ning1,2,3, Yulu Peng1,2,3, Chunping Yu1,2,3, Ya Ding2,3,5, Desheng Weng2,3,5, Jianchuan Xia2,3,5, Lijuan Jiang1,2,3, Shengjie Guo1,2,3, Hui Han1,2,3, Fangjian Zhou1,2,3, Pei Dong1,2,3 1Department of Urology, Sun Yat-sen University Cancer Center, Guangzhou, China; 2State Key Laboratory of Oncology in Southern China, Guangzhou, China; 3Collaborative Innovation Center for Cancer Medicine, Guangzhou, China; 4Department of Radiology, Sun Yat-sen University Cancer Center, Guangzhou, China; 5Department of Biotherapy, Sun Yat-sen University Cancer Center, Guangzhou, China Contributions: (I) Conception and design: Z Zhang, P Dong, F Zhou; (II) Administrative support: H Han, J Xia, D Weng; (III) Provision of study materials or patients: H Liu, K Ning, Y Peng, C Yu, Y Ding; (IV) Collection and assembly of data: Z Zhang, L Xiong, Z Wu, L Jiang, S Guo; (V) Data analysis and interpretation: Z Zhang, L Xiong, Z Wu; (VI) Manuscript writing: All authors; (VII) Final approval of manuscript: All authors. #These authors contributed equally to this work. Correspondence to: Zhiling Zhang; Pei Dong. Department of Urology, Sun Yat-sen University Cancer Center, 651, Dong Feng Dong Road, Guangzhou 510060, China. Email: [email protected]; [email protected]. Background: Radical/cytoreductive nephrectomy or nephron-sparing surgery may be thought to be not safe or unfeasible in some renal cell carcinoma (RCC) patients in which tumor is locally advanced or highly complicated. -

Tyrosine Kinase Inhibitors in Cancer: Breakthrough and Challenges of Targeted Therapy
cancers Review Tyrosine Kinase Inhibitors in Cancer: Breakthrough and Challenges of Targeted Therapy 1,2, 3,4 1 2 3, Charles Pottier * , Margaux Fresnais , Marie Gilon , Guy Jérusalem ,Rémi Longuespée y 1, and Nor Eddine Sounni y 1 Laboratory of Tumor and Development Biology, GIGA-Cancer and GIGA-I3, GIGA-Research, University Hospital of Liège, 4000 Liège, Belgium; [email protected] (M.G.); [email protected] (N.E.S.) 2 Department of Medical Oncology, University Hospital of Liège, 4000 Liège, Belgium; [email protected] 3 Department of Clinical Pharmacology and Pharmacoepidemiology, University Hospital of Heidelberg, 69120 Heidelberg, Germany; [email protected] (M.F.); [email protected] (R.L.) 4 German Cancer Consortium (DKTK)-German Cancer Research Center (DKFZ), 69120 Heidelberg, Germany * Correspondence: [email protected] Equivalent contribution. y Received: 17 January 2020; Accepted: 16 March 2020; Published: 20 March 2020 Abstract: Receptor tyrosine kinases (RTKs) are key regulatory signaling proteins governing cancer cell growth and metastasis. During the last two decades, several molecules targeting RTKs were used in oncology as a first or second line therapy in different types of cancer. However, their effectiveness is limited by the appearance of resistance or adverse effects. In this review, we summarize the main features of RTKs and their inhibitors (RTKIs), their current use in oncology, and mechanisms of resistance. We also describe the technological advances of artificial intelligence, chemoproteomics, and microfluidics in elaborating powerful strategies that could be used in providing more efficient and selective small molecules inhibitors of RTKs. -

202324Orig1s000
CENTER FOR DRUG EVALUATION AND RESEARCH APPLICATION NUMBER: 202324Orig1s000 CLINICAL PHARMACOLOGY AND BIOPHARMACEUTICS REVIEW(S) RECOMMENDATION: 1. The proposed film coat weight gain (b) (4) is acceptable for the 1 mg and 5 mg tablet. • This specification is supported by the information received on December 21, 2011 and on the bioequivalency between both strengths, the formulation characteristics, and the in vitro dissolution performance. Kareen Riviere, PhD Sandra Suarez Sharp, PhD Biopharmaceutics Reviewer Senior Biopharmaceutics Reviewer Office of New Drugs Quality Assessment Office of New Drugs Quality Assessment cc: Angelica Dorantes, Ph.D. Drug Product Film Coating Weight The Applicant proposed a film coat weight (b) (4) to control the weight of film coating applied to tablets. However, there no data was submitted supporting this proposed film coat specification. The following Biopharmaceutics comment related to film coating specification was made in the Information Request Letter dated September 26, 2011. FDA Query 36 Indicate if (b) (4) % of film coat has any impact on dissolution with supporting data, if available. Sponsor Response (b) (4) 4 Page(s) has been Withheld in Full as B4 (CCI/ TS) immediately following this 2 Reference ID: 3076627 --------------------------------------------------------------------------------------------------------- This is a representation of an electronic record that was signed electronically and this page is the manifestation of the electronic signature. --------------------------------------------------------------------------------------------------------- -

Avelumab in Combination with Axitinib Advanced Renal Cell Carcinoma
THE CLATTERBRIDGE CANCER CENTRE NHS FOUNDATION TRUST Systemic Anti Cancer Therapy Protocol Avelumab in Combination with Axitinib Advanced Renal Cell Carcinoma PROTOCOL REF: MPHAAAAUR (Version No: 1.0) This protocol has been temporarily amended - please see the SRG Guidelines during COVID-19 Urology Cancer. Approved for use in: First line treatment for adult patients with advanced renal cell carcinoma which has either a clear cell component or is one of the types of RCC outlined in blueteq the form. ECOG performance status 0 or 1. *Blueteq registration required: check Blueteq for full eligibility criteria* Dosage: Drug Dose Route Frequency Avelumab 800mg IV infusion Days 1 and 15 5mg twice Axitinib Oral Every 28 days daily Every 28 days until disease progression or unacceptable toxicity Administration: Take axitinib with or without food approximately 12 hours apart, swallow whole with a glass of water. Emetogenic risk: Mildly emetogenic Supportive treatments: Domperidone- 10mg three times a day if required Issue Date: 11th December 2020 Page 1 of 10 Protocol reference: MPHAAAAUR Review Date: December 2023 Author: Anna Burke Authorised by: Drug & Therapeutics Committee Version No: 1.0 THE CLATTERBRIDGE CANCER CENTRE NHS FOUNDATION TRUST Extravasation risk: Avelumab (monoclonal antibody) - neutral. Refer to Clatterbridge Policy ‘Prevention and Management of Extravasation Injuries’ for further guidance. Dosing in renal and hepatic impairment: GFR ≥ 30 ml/min: no dose adjustment is needed GFR <30 ml/min and patients on haemodialysis: no need for Avelumab dose adjustment is expected. Use with caution as not tested Renal in this patient cohort Renal impairment: no dose adjustment is needed Axitinib Haemodialysis no need for dose adjustment is expected Mild: Bil >1.0-1.5 x ULN and any AST or Bil ≤ULN and AST >ULN no dose adjustment is needed Avelumab Moderate: Bil 1.5-3 x ULN, with any AST or severe Bil >3.0-10 x ULN, with any AST no need for dose adjustment is expected. -

(Vismodegib) for Gleevec
TOOLS APPROVED DRUGS The recommended dose of imatinib or mildly impaired renal function because for adjuvant treatment is 400 mg/day of the potential risk of subtherapeutic • Genentech (www.gene.com), a member administered with meals daily for three exposure to methotrexate. of the Roche Group, announced the years. The optimal duration of treatment Food and Drug Administration (FDA) has is not known. • Millennium: The Takeda Oncol- approved Erivedge™ (vismodegib) ogy Company (www.millennium.com) capsules for the treatment of adults • The FDA approved Inlyta (axitinib announced that the FDA has approved with metastatic basal cell carcinoma tablets) (Pfizer, Inc. www.pfizer. a supplemental new drug application (BCC) or with locally advanced BCC that com), a kinase inhibitor, for treatment for Velcade® (bortezomib), which has recurred following surgery or who are of patients with advanced renal cell updates the label to include the subcu- not candidates for surgery, and who are carcinoma (RCC) after failure of one prior taneous method of administration in all not candidates for radiation. systemic therapy. The approval is based approved indications: multiple myeloma Vismodegib inhibits the Hedge- on data from the Phase III AXIS trial, and mantel cell lymphoma after at least hog pathway, an important embryonic which demonstrated that Inlyta signifi- one prior therapy. developmental pathway. Reproductive cantly extended progression free survival toxicology studies in rats demonstrated (PFS) [HR=0.67, 0.54-0.81, P<0.0001] that vismodegib exposure during organo- with a median PFS of 6.7 months (95% DRUGS IN THE NEWS genesis results in embryo-fetal death at CI: 6.3,8.6) compared with 4.7 months higher exposures and severe birth defects (95% CI: 4.6,5.6) for those treated with • Pinnacle Biologics, Inc.