Frontoparietal Bone in Extinct Palaeobatrachidae (Anura): Its Variation and Taxonomic Value
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Biostratigraphy and Paleoecology of Continental Tertiary Vertebrate Faunas in the Lower Rhine Embayment (NW-Germany)
Netherlands Journal of Geosciences / Geologie en Mijnbouw 81 (2): 177-183 (2002) Biostratigraphy and paleoecology of continental Tertiary vertebrate faunas in the Lower Rhine Embayment (NW-Germany) Th. Mors Naturhistoriska Riksmuseet/Swedish Museum of Natural History, Department of Palaeozoology, P.O. Box 50007, SE-104 05 Stockholm, Sweden; e-mail: [email protected] Manuscript received: October 2000; accepted: January 2002 ^ Abstract This paper discusses the faunal content, the mammal biostratigraphy, and the environmental ecology of three important con tinental Tertiary vertebrate faunas from the Lower Rhine Embayment. The sites investigated are Rott (MP 30, Late Oligocene), Hambach 6C (MN 5, Middle Miocene), Frechen and Hambach 11 (both MN 16, Late Pliocene). Comparative analysis of the entire faunas shows the assemblages to exhibit many conformities in their general composition, presumably re sulting from their preference for wet lowlands. It appears that very similar environmental conditions for vertebrates reoc- curred during at least 20 Ma although the sites are located in a tectonically active region with high subsidence rates. Differ ences in the faunal composition are partly due to local differences in the depositional environment of the sites: lake deposits at the margin of the embayment (Rott), coal swamp and estuarine conditions in the centre of the embayment (Hambach 6C), and flood plain environments with small rivulets (Frechen and Hambach 1 l).The composition of the faunal assemblages (di versity and taxonomy) also documents faunal turnovers with extinctions and immigrations (Oligocene/Miocene and post- Middle Miocene), as a result of changing climate conditions. Additional vertebrate faunal data were retrieved from two new assemblages collected from younger strata at the Hambach mine (Hambach 11C and 14). -

Baez NHM 1997.Pdf (3.622Mb)
HERP QL 668 B33 l':iif;,O0i.U'.Z06l("'^ ocxentxpc Papers Natural History Museum The University of Kansas 29 October 1997 Number 4:1^1 Redescription of the Paleogene Shelania pascuali from Patagonia and Its Bearing on the Relationships of Fossil and Recent Pipoid Frogs >. By o O Ana Maria BAez^ and Linda Trueb- o ^Departamento de Geologia, FacuUad de Ciencias Exactas, Universidad de 0) w >. > t- Buenos Aires, Pabellon II, Ciudad Universitaria, 1428 Buenos Aires, Argentina CO !-> >^ ^ £ -^ '^Division of Herpetology, Natural Histoiy Museum, and Department of P Kansas 66045-2454, J3 o, Systematics and Ecology, The University of Kansas, Lawrence, USA a o t3 o CO ^ t. CONTENTS ° CO I ABSTRACT 2 « RESUMEN 2 5S INTRODUCTION 2 Previous Paleontological Work 4 Acknowledgments 4 MATERIALS AND METHODS 5 General Methodology 5 Cladistic Methodology 5 Specimens Examined 6 STRATIGRAPHIC PROVENANCE AND AGE OF MATERIAL 6 REDESCRIPTION OF SHELANIA 8 ANALYSIS OF CHARACTERS 16 RESULTS 31 DISCUSSION 35 Taxonomic Considerations 35 Characters 36 LITERATURE CITED 37 APPENDIX 40 © Natural History Museum, The University of Kansas ISSN No. 1094-0782 — 2 Scientific Papers, Natural History Museum, The University of Kansas 1960, is redescribed on the basis of a series of 30 recently I ABSTIMCT Shdania pascuali Casamiquela, .'-\ ' . discovered specimens, which range in estimated snout-vent length from 30-100 mm, from the Paleo- y ' gene of Patagonia. This large pipoid anuran is distinguished by possessing a long, narrow braincase; an hourglass-shaped frontoparietal; a robust antorbital process on the edentate maxilla; long, straight {/) '/^ I ilia that describe a V-shape in dorsal profile; and a trunk that is long relative to the lengths of the head and limbs. -

A Review of Neusibatrachus Wilferti, an Early Cretaceous Frog from the Montsec Range, Northeastern Spain
A review of Neusibatrachus wilferti, an Early Cretaceous frog from the Montsec Range, northeastern Spain ANA M. BÁEZ and BORJA SANCHIZ Báez, A.M. and Sanchiz, B. 2007. A review of Neusibatrachus wilferti, an Early Cretaceous frog from the Montsec Range, northeastern Spain. Acta Palaeontologica Polonica 52 (3): 477–487. Neusibatrachus wilferti is an anuran from the late Berriasian–early Valanginian fossiliferous lacustrine limestones that are exposed in the eastern part of the Montsec Range, province of Lleida, Spain. It was originally described by Seiffert in 1972 and its phylogenetic position has since been discussed. Neusibatrachus has been considered an undeterminable fos− sil, an abnormal individual, or a primitive palaeobatrachid. Here we redescribe the only available specimen, and clarify features, such as absence of palatines, nine presacrals, and procoelous vertebral centra, that have been the subject of previ− ous debates. We consider the specimen to be a postmetamorphic individual and make developmental interpretations of some of its characters. In particular, we provide evidence of a living anuran (Rana iberica) that resembles Neusibatrachus in the development of intervertebral articulations. Neusibatrachus is considered a valid genus, which differs from other anurans, except for the pipoids, in the joint presence of an azygous frontoparietal and a parasphenoid lacking the subotic alae, although it differs from the pipoids in having nine presacral vertebrae. Morphological evidence indicates that Neusi− batrachus is related to Xenoanura, the pipoid branch in the living Amphibia Tree of Life based on molecular data. More− over, it might be a member of the pipoid clade proper, which presently includes the Pipidae, Rhinophrynidae, and several fossil taxa, including the Palaeobatrachidae, although the evidence is not conclusive. -

Exceptional Fossil Preservation During CO2 Greenhouse Crises? Gregory J
Palaeogeography, Palaeoclimatology, Palaeoecology 307 (2011) 59–74 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Exceptional fossil preservation during CO2 greenhouse crises? Gregory J. Retallack Department of Geological Sciences, University of Oregon, Eugene, Oregon 97403, USA article info abstract Article history: Exceptional fossil preservation may require not only exceptional places, but exceptional times, as demonstrated Received 27 October 2010 here by two distinct types of analysis. First, irregular stratigraphic spacing of horizons yielding articulated Triassic Received in revised form 19 April 2011 fishes and Cambrian trilobites is highly correlated in sequences in different parts of the world, as if there were Accepted 21 April 2011 short temporal intervals of exceptional preservation globally. Second, compilations of ages of well-dated fossil Available online 30 April 2011 localities show spikes of abundance which coincide with stage boundaries, mass extinctions, oceanic anoxic events, carbon isotope anomalies, spikes of high atmospheric carbon dioxide, and transient warm-wet Keywords: Lagerstatten paleoclimates. Exceptional fossil preservation may have been promoted during unusual times, comparable with fi Fossil preservation the present: CO2 greenhouse crises of expanding marine dead zones, oceanic acidi cation, coral bleaching, Trilobite wetland eutrophication, sea level rise, ice-cap melting, and biotic invasions. Fish © 2011 Elsevier B.V. All rights reserved. Carbon dioxide Greenhouse 1. Introduction Zeigler, 1992), sperm (Nishida et al., 2003), nuclei (Gould, 1971)and starch granules (Baxter, 1964). Taphonomic studies of such fossils have Commercial fossil collectors continue to produce beautifully pre- emphasized special places where fossils are exceptionally preserved pared, fully articulated, complex fossils of scientific(Simmons et al., (Martin, 1999; Bottjer et al., 2002). -
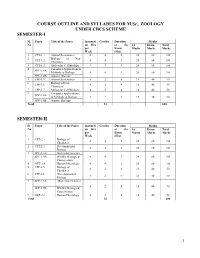
COURSE OUTLINE and SYLLABUS for M.Sc., ZOOLOGY UNDER CBCS SCHEME SEMESTER-I
COURSE OUTLINE AND SYLLABUS FOR M.Sc., ZOOLOGY UNDER CBCS SCHEME SEMESTER-I Sl. Paper Title of the Paper Instructi Credits Duration Marks No on Hrs of the IA Exam Total per Exam Marks Marks Marks Week (Hrs) 1 CPT-1.1 Animal Systematics 4 4 3 20 80 100 2 Biology of Non- CPT-1.2 4 4 3 20 80 100 Chordates 3 CPT-1.3 Molecular Cell Biology 4 4 3 20 80 100 4 Computer Applications & SPT-1.4A Methods in Biology 4 4 3 20 80 100 SPT-1.4B Aquatic Biology 5 CPP-1.5 Animal Systematics 4 2 4 10 40 50 6 Biology of Non- CPP-1.6 4 2 4 10 40 50 Chordates 7 CPP-1.7 Molecular Cell Biology 4 2 4 10 40 50 8 Computer Applications SPP-1.8A & Methods in Biology 4 2 4 10 40 50 SPP-1.8B Aquatic Biology Total 24 600 SEMESTER-II Sl. Paper Title of the Paper Instructi Credits Duration Marks No on Hrs of the IA Exam Total per Exam Marks Marks Marks Week (Hrs) 1 CPT-2.1 Biology of 4 4 3 20 80 100 Chordates 2 CPT-2.2 Developmental 4 4 3 20 80 100 Biology 3 SPT-2.3A Molecular Genetics SPT-2.3B Wildlife Biology & 4 4 3 20 80 100 Conservation 4 OET-2.4 Human Physiology 4 4 3 20 80 100 5 CPP-2.5 Biology of 4 2 4 10 40 50 Chordates 6 CPP-2.6 Developmental 4 2 4 10 40 50 Biology 7 SPP-2.7A Molecular Genetics 4 2 4 10 40 50 SPP-2.7B Wildlife Biology & Conservation 8 OEP-2.8 Human Physiology 4 2 4 10 40 50 Total 24 600 1 SEMESTER-III Sl. -

ROCEK, Z. and WUTTKE, M. (2010) Amphibia of Enspel (Late
Palaeobio Palaeoenv (2010) 90:321–340 DOI 10.1007/s12549-010-0042-0 ORIGINAL PAPER Amphibia of Enspel (Late Oligocene, Germany) ZbyněkRoček & Michael Wuttke Received: 23 April 2010 /Revised: 9 July 2010 /Accepted: 12 August 2010 /Published online: 29 September 2010 # Senckenberg Gesellschaft für Naturforschung and Springer 2010 Abstract Amphibia from the Late Oligocene (MP 28) One specimen is a large premetamorphic tadpole (no locality Enspel, Germany are represented by two caudates: rudimentary limbs) with a total body length of 147 mm. a hyperossified salamandrid Chelotriton paradoxus and an Anatomically, it can be equally assigned to Pelobates or to indeterminate salamandrid different from Chelotriton in Eopelobates; the second possibility was excluded only on proportions of vertebral column. Anurans are represented the basis of absence of adult Eopelobates in this locality. by two forms of the genus Palaeobatrachus, one of which is nearly as large as P. gigas (now synonymized with P. Keywords Enspel . Oligocene . Salamandridae . grandipes). Pelobates cf. decheni, represented in this Chelotriton . Anura . Palaeobatrachus . Pelobates . Rana locality by three nearly complete adult skeletons and a large number of tadpoles, is the earliest record for the Abbreviation genus. Compared with later representatives of the genus, it DP FNSP Department of Palaeontology Faculty of does not yet possess specializations for burrowing. Ranidae Natural Sciences, Prague are represented by two rather fragmentary and incomplete skeletons referred to as Rana sp. A comparatively large series of tadpoles was assigned to the Pelobatidae on the basis of tripartite frontoparietal complex. Most of them are Introduction premetamorphic larvae, and a few older ones are post- metamorphic, but they do not exceed Gossner stage 42. -

Larval Development in Oligocene Palaeobatrachid Frogs
Larval development in Oligocene palaeobatrachid frogs ZBYNĚK ROČEK Roček, Z. 2003. Larval development in Oligocene palaeobatrachid frogs. Acta Palaeontologica Polonica 48 (4): 595–607. A detailed account of the development of skeletal and some soft−tissue structures is based on 171 fossil tadpoles and meta− morphosing froglets of Palaeobatrachus sp. from the Late Oligocene of the Czech Republic (locality Bechlejovice). Their exceptionally good preservation resulted from fossilization in diatomites. The fossil developmental series was com− pared with normal development of the contemporary anuran Xenopus laevis (Pipidae) represented by cleared and stained (alizarin/toluidin−blue) whole−mount specimens. The comparison revealed that in spite of differences in the sequence of ossification and its timing (e.g., ossification of the otic capsules and ribs was retarded in Xenopus whereas dermal ossifi− cation was retarded in Palaeobatrachus), in the number of free ribs, and in composition of the sacral region (the synsacrum in Palaeobatrachus involves two posterior presacrals, whereas there is a single sacral in Xenopus), both gen− era were similar in great number of anatomical features that appear during development. The most important difference is the shape of vertebral centrum (procoelous in Palaeobatrachus, opisthocoelous in all Pipidae) which is formed in com− paratively early developmental stages. A view that could result from anatomical comparisons is that Palaeobatrachus could be derived from the Pipidae, but this is doubtful due to biostratigraphic and palaeogeographic discrepancies. The earliest palaeobatrachids were recorded from the Late Cretaceous of Europe but pipids could not invade northern conti− nents after the Early Cretaceous when the Tethys Sea prevented interchanges of anuran faunas. -

Chondrocranial and Oral Morphology of Pipoid Frogs Charles Christopher Swart
University of Richmond UR Scholarship Repository Master's Theses Student Research 1998 Chondrocranial and oral morphology of pipoid frogs Charles Christopher Swart Follow this and additional works at: http://scholarship.richmond.edu/masters-theses Part of the Biology Commons Recommended Citation Swart, Charles Christopher, "Chondrocranial and oral morphology of pipoid frogs" (1998). Master's Theses. Paper 1065. This Thesis is brought to you for free and open access by the Student Research at UR Scholarship Repository. It has been accepted for inclusion in Master's Theses by an authorized administrator of UR Scholarship Repository. For more information, please contact [email protected]. Chondrocranial and Oral Morphology of Pipoid Frogs by Charles Christopher Swart I certify that I have read this thesis and find that, in scope and quality, it satisfies the requirements for the degree of Master of Science. Dr.Joi<rlttayden Dr.G~;?j_~ T Other Faculty Members: TT Chondrocranial and Oral Morphology of Pipoid Frogs Charles Christopher Swart Masters of Science in Biology University of Richmond, 1998 Thesis Director: Rafael 0. de Sa Abstract. The Pipoidea are a diverse group of frogs. Their diversity is demonstrated in their morphology, ecology, and behavior. One pipoid species, Xenopus laevis, has been used as a model system of developmental, physiological, and molecular studies of vertebrates. My work has focused on the developmental morphology of the chondrocranium and oral morphology of four pipoid taxa: Hymenochirus boettgeri, Rhinophrynus dorsalis, Pipa carvalhoi, and Xenopus laevis. Previous studies have suggested that the Anura may be diphyletic based on the unique characteristics of the chondrocanial morphology of pipoids. -

Neogene Amphibians and Reptiles (Caudata, Anura, Gekkota, Lacertilia, and Testudines) from the South of Western Siberia, Russia, and Northeastern Kazakhstan
Neogene amphibians and reptiles (Caudata, Anura, Gekkota, Lacertilia, and Testudines) from the south of Western Siberia, Russia, and Northeastern Kazakhstan Davit Vasilyan1,2,3, Vladimir S. Zazhigin4 and Madelaine Bo¨hme1,5 1 Department of Geosciences, Eberhard Karls University Tu¨bingen, Tu¨bingen, Germany 2 JURASSICA Museum, Porrentruy, Switzerland 3 Department of Geosciences, University of Fribourg, Fribourg, Switzerland 4 Institute of Geology, Russian Academy of Sciences, Moscow, Russia 5 Senckenberg Center for Human Evolution and Palaeoecology, Eberhard Karls University Tu¨bingen, Tu¨bingen, Germany ABSTRACT Background: The present-day amphibian and reptile fauna of Western Siberia are the least diverse of the Palaearctic Realm, as a consequence of the unfavourable climatic conditions that predominate in this region. The origin and emergence of these herpetofaunal groups are poorly understood. Aside from the better-explored European Neogene localities yielding amphibian and reptile fossil remains, the Neogene herpetofauna of Western Asia is understudied. The few available data need critical reviews and new interpretations, taking into account the more recent records of the European herpetofauna. The comparison of this previous data with that of European fossil records would provide data on palaeobiogeographic affiliations of the region as well as on the origin and emergence of the present-day fauna of Western Siberia. An overview of the earliest occurrences of certain amphibian lineages is still needed. In addition, studies that address such knowledge gaps can be Submitted 16 August 2016 useful for molecular biologists in their calibration of molecular clocks. Accepted 24 January 2017 Methods and Results: In this study, we considered critically reviewed available data Published 23 March 2017 from amphibian and reptile fauna from over 40 Western Siberian, Russian and Corresponding author Northeastern Kazakhstan localities, ranging from the Middle Miocene to Early Davit Vasilyan, [email protected] Pleistocene. -

Retallack 2011 Lagerstatten
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright Author's personal copy Palaeogeography, Palaeoclimatology, Palaeoecology 307 (2011) 59–74 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Exceptional fossil preservation during CO2 greenhouse crises? Gregory J. Retallack Department of Geological Sciences, University of Oregon, Eugene, Oregon 97403, USA article info abstract Article history: Exceptional fossil preservation may require not only exceptional places, but exceptional times, as demonstrated Received 27 October 2010 here by two distinct types of analysis. First, irregular stratigraphic spacing of horizons yielding articulated Triassic Received in revised form 19 April 2011 fishes and Cambrian trilobites is highly correlated in sequences in different parts of the world, as if there were Accepted 21 April 2011 short temporal intervals of exceptional preservation globally. Second, compilations of ages of well-dated fossil Available online 30 April 2011 localities show spikes of abundance which coincide with stage boundaries, mass extinctions, oceanic anoxic events, carbon isotope anomalies, spikes of high atmospheric carbon dioxide, and transient warm-wet Keywords: Lagerstatten paleoclimates. -

Icp Publications 2006-2021
ICP PUBLICATIONS 2006-2021 Last updated: 1 September 2021 In press and published online _______________________________________________________________________ SCI papers (indexed in JCR) 1. Abella, J., Martín-Perea, D. M., Valenciano, A., Hontecillas, D., Montoya, P., & Morales, J. (2021, published online). Coprolites in natural traps: direct evidence of bone eating carnivorans from the Late Miocene site of Batallones-3 (Madrid, Spain). Lethaia. https://doi.org/10.1111/let.12438 2. Agustí, J., Espresate, J., & Piñero, P. (2020, published in press). Dental variation in the endemic dormouse Hypnomys Bate 1918 and its implications for the palaeogeographic evolution of the Balearic Islands (Western Mediterranean) during the late Neogene-Quaternary. Historical Biology. https://doi.org/10.1080/08912963.2020.1852557 3. Alba, D. M., Robles, J. M., Valenciano, A., Abella, J., & Casanovas-Vilar, I. (2021, published online). A new species of Eomellivora from the latest Aragonian of Abocador de Can Mata (NE Iberian Peninsula). Historical Biology. https://doi.org/10.1080/08912963.2021.1943380 4. Arias-Martorell, J., Zeininger, A., & Kivell, T. L. (in press). Trabecular structure of the elbow reveals divergence in knuckle-walking biomechanical strategies of African apes. Evolution. 5. Bouchet, F., Urciuoli, A., Beaudet, A., Pina, M., Moyà-Solà, S., & Alba, D. M. (in press). Comparative anatomy of the carotid canal in the Miocene small-bodied catarrhine Pliobates cataloniae. Journal of Human Evolution. 6. Caballero, Ó., Montoya, P., Crespo, V. D., Morales, J., & Abella, J. (2020, published online). The autopodial skeleton of Paracamelus aguirrei (Morales 1984) (Tylopoda, Mammalia) from the late Miocene site of Venta del Moro (Valencia, Spain). Journal of Iberian Geology. -

The Present Paper Describes for the First Time in Detail the Herpeto Fauna from the Locality of W~Ze II
Acta Palaeontologica Polonica Vol. 29, No . 3-4 pp.209-226 VVarszavva, 1984 MARIAN MLYNARSKI, ZBIGNIEVV SZYNDLAR, RICHARD \.ESTES and BORJA SANCHlz AMPHIBIANS AND REPTILES FROM THE PLIOCENE LOCALITY OF WEiZE II NEAR DZIALOSZYN (POLAND) . MLYNARSKI, M., SZYNDLAR, Z. , ESTES, R. and SANCHIZ, B.: Amphibians and reptiles from the Pliocene locality of VV<;ze II near Dzialoszyn (Poland). Acta Palaeont. Polonica, 29, 3-4, 209-226, 1985 (1984). , The following forms have been recognized in the Upper Pliocene fauna from the karstic deposits near Dzialoszyn: Mtoproteus wezet sp, n. (Proteidae), pataeoba trachus sp. and p!tobatrachus cr. tanghae (Palaeobatrachidae), Petobates fuscus (Pelobatidae), Bufo bUfo (Bufonidae), Rana sp, (Ranidae), Emys orbtcutarts an ttgua (Emydidae), Ophlsaurus pannontcus and Anguts ct. fraglUs (Anguidae), La certa cr. v trtdts and Lacerta sp, (Lacertidae), Etaphe paratongtsstma and Natrtx ct. tongtvertebrata (Colubridae). The majority of the recognized taxa, including living forms (e.g, Bufo tniio, Emys orblcutarts, Anguts fraglUs) as well as extinct ones (Pltobatrachus tanqhae. Natrtx tongtvertebrata), were widespread in the Euro pean Neogene lind ·(e ven tuall y) Pleistocene. Key W 0 r d s : Amphibia, Reptilia, Pliocene, Poland. Marian ilnynarski and Zbtgniew Szyndtar. Zaklad zootoglt Systematycznej t 00 Awtadczatnej, Potska Akademia Nauk, S!awkowska 17, 31-016 Krak6w, Potand. Richard Estes, Department of zootogy, CoUege Of Sctence, San otego, CA 92182-0063, USA. Borja Sanchtz, Museo Nactonat de Ctenctas Naturates (C.S.I.C.), Castettana, 80, Madrid-G, Spa tn. INTRODUCTION The present paper describes for the first time in detail the herpeto fauna from the locality of W~ze II. This fossil site was discovered by Sulimski (1962) and named W~ze II in order to distinguish it from the nearby locality W~Ze 1.