ROCEK, Z. and WUTTKE, M. (2010) Amphibia of Enspel (Late
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Biostratigraphy and Paleoecology of Continental Tertiary Vertebrate Faunas in the Lower Rhine Embayment (NW-Germany)
Netherlands Journal of Geosciences / Geologie en Mijnbouw 81 (2): 177-183 (2002) Biostratigraphy and paleoecology of continental Tertiary vertebrate faunas in the Lower Rhine Embayment (NW-Germany) Th. Mors Naturhistoriska Riksmuseet/Swedish Museum of Natural History, Department of Palaeozoology, P.O. Box 50007, SE-104 05 Stockholm, Sweden; e-mail: [email protected] Manuscript received: October 2000; accepted: January 2002 ^ Abstract This paper discusses the faunal content, the mammal biostratigraphy, and the environmental ecology of three important con tinental Tertiary vertebrate faunas from the Lower Rhine Embayment. The sites investigated are Rott (MP 30, Late Oligocene), Hambach 6C (MN 5, Middle Miocene), Frechen and Hambach 11 (both MN 16, Late Pliocene). Comparative analysis of the entire faunas shows the assemblages to exhibit many conformities in their general composition, presumably re sulting from their preference for wet lowlands. It appears that very similar environmental conditions for vertebrates reoc- curred during at least 20 Ma although the sites are located in a tectonically active region with high subsidence rates. Differ ences in the faunal composition are partly due to local differences in the depositional environment of the sites: lake deposits at the margin of the embayment (Rott), coal swamp and estuarine conditions in the centre of the embayment (Hambach 6C), and flood plain environments with small rivulets (Frechen and Hambach 1 l).The composition of the faunal assemblages (di versity and taxonomy) also documents faunal turnovers with extinctions and immigrations (Oligocene/Miocene and post- Middle Miocene), as a result of changing climate conditions. Additional vertebrate faunal data were retrieved from two new assemblages collected from younger strata at the Hambach mine (Hambach 11C and 14). -

The World at the Time of Messel: Conference Volume
T. Lehmann & S.F.K. Schaal (eds) The World at the Time of Messel - Conference Volume Time at the The World The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment and the History of Early Primates 22nd International Senckenberg Conference 2011 Frankfurt am Main, 15th - 19th November 2011 ISBN 978-3-929907-86-5 Conference Volume SENCKENBERG Gesellschaft für Naturforschung THOMAS LEHMANN & STEPHAN F.K. SCHAAL (eds) The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates 22nd International Senckenberg Conference Frankfurt am Main, 15th – 19th November 2011 Conference Volume Senckenberg Gesellschaft für Naturforschung IMPRINT The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates 22nd International Senckenberg Conference 15th – 19th November 2011, Frankfurt am Main, Germany Conference Volume Publisher PROF. DR. DR. H.C. VOLKER MOSBRUGGER Senckenberg Gesellschaft für Naturforschung Senckenberganlage 25, 60325 Frankfurt am Main, Germany Editors DR. THOMAS LEHMANN & DR. STEPHAN F.K. SCHAAL Senckenberg Research Institute and Natural History Museum Frankfurt Senckenberganlage 25, 60325 Frankfurt am Main, Germany [email protected]; [email protected] Language editors JOSEPH E.B. HOGAN & DR. KRISTER T. SMITH Layout JULIANE EBERHARDT & ANIKA VOGEL Cover Illustration EVELINE JUNQUEIRA Print Rhein-Main-Geschäftsdrucke, Hofheim-Wallau, Germany Citation LEHMANN, T. & SCHAAL, S.F.K. (eds) (2011). The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates. 22nd International Senckenberg Conference. 15th – 19th November 2011, Frankfurt am Main. Conference Volume. Senckenberg Gesellschaft für Naturforschung, Frankfurt am Main. pp. 203. -

Frogs (Amphibia, Anura) from the Eocene and Oligocene of the Phosphorites Du Quercy (France)
FOSSIL IMPRINT • vol. 72 • 2016 • no. 1-2 • pp. 53–66 (formerly ACTA MUSEI NATIONALIS PRAGAE, Series B – Historia Naturalis) FROGS (AMPHIBIA, ANURA) FROM THE EOCENE AND OLIGOCENE OF THE PHOSPHORITES DU QUERCY (FRANCE). AN OVERVIEW JEAN-CLAUDE RAGE Sorbonne Universités, CR2P CNRS-MNHN-UPMC Paris 6, Département Histoire de la Terre, Muséum National d’Histoire Naturelle, CP 38, 57 rue Cuvier, 75005 Paris, France; e-mail: [email protected]. Rage, J.-C. (2016): Frogs (Amphibia, Anura) from the Eocene and Oligocene of the Phosphorites du Quercy (France). An overview. – Fossil Imprint, 72(1-2): 53–66, Praha. ISSN 2533-4050 (print), ISSN 2533-4069 (on-line). Abstract: Anuran assemblages from the Eocene and Oligocene of the Phosphorites du Quercy (southwestern France) are documented by a modest number of isolated bones, incomplete skulls, and some ‘mummies’. However, at the family level, the diversity is not significantly lower than in coeval frog assemblages from other regions. By contrast, a larger number of amniote specimens and taxa are known from the Phosphorites. The sparse anuran record within the Phosphorites most likely results from the karstic environment in which the fossiliferous sites formed. Such an environment is not favorable for animals dependent on water and moisture. Upper middle and upper Eocene localities in the Phosphorites produced Alytidae, Pelobatidae, Pelodytidae and ranoids. The presence of bufonids or microhylids, and rhacophorids cannot be definitely rejected, and potentially distinctive, but unidentifiable taxa also may be present. The occurrence of Pelodytidae in the Eocene of Europe is confirmed by diagnostic bones from the Phosphorites. In Oligocene localities in the Phosphorites, anurans are even less diverse, with only Alytidae, Pelobatidae and ranoids having been recovered. -

Strange Tadpoles from the Lower Miocene of Turkey: Is Paedogenesis Possible in Anurans?
Strange tadpoles from the lower Miocene of Turkey: Is paedogenesis possible in anurans? ALAIN DUBOIS, STÉPHANE GROSJEAN, and JEAN−CLAUDE PAICHELER Dubois, A., Grosjean, S., and Paicheler, J.−C. 2010. Strange tadpoles from the lower Miocene of Turkey: Is paedogenesis possible in anurans? Acta Palaeontologica Polonica 55 (1): 43–55. Fossil material from the lower Miocene collected in the basin lake of Beşkonak (Turkey) included 19 slabs showing 19 amphibian anuran tadpoles of rather large size, at Gosner stages 36–38. These well preserved specimens show many mor− phological and skeletal characters. They are here tentatively referred to the genus Pelobates. Two of these tadpoles show an unusual group of black roundish spots in the abdominal region, and a third similar group of spots is present in another slab but we were unable to state if it was associated with a tadpole or not. Several hypotheses can be proposed to account for these structures: artefacts; intestinal content (seeds; inert, bacterial or fungal aggregations; eggs); internal or external parasites; diseases; eggs produced by the tadpole. The latter hypothesis is discussed in detail and is shown to be unlikely for several reasons. However, in the improbable case where these spots would correspond to eggs, this would be the first reported case of natural paedogenesis in anurans, a phenomenon which has been so far considered impossible mostly for anatomical reasons (e.g., absence of space in the abdominal cavity). Key words: Amphibia, Anura, egg, fossil, paedogenesis, tadpole, Miocene, Turkey. Alain Dubois [[email protected]] and Stéphane Grosjean [[email protected]], Département de Sytématique & Evolu− tion, Muséum National d’Histoire Naturelle, UMR 7205 Origine, Structure et Evolution de la Biodiversité, Reptiles et Amphibiens, Case 30, 25 rue Cuvier, 75005 Paris, France; Jean−Claude Paicheler [[email protected]], Laboratoire de Paléoparasitologie, UFR de Pharmacie, 51 rue Cognacq Jay, 51096 Reims Cedex, France. -

Curriculum Vitae
CURRICULUM VITAE AMY C. HENRICI Collection Manager Section of Vertebrate Paleontology Carnegie Museum of Natural History 4400 Forbes Avenue Pittsburgh, Pennsylvania 15213-4080, USA Phone:(412)622-1915 Email: [email protected] BACKGROUND Birthdate: 24 September 1957. Birthplace: Pittsburgh. Citizenship: USA. EDUCATION B.A. 1979, Hiram College, Ohio (Biology) M.S. 1989, University of Pittsburgh, Pennsylvania (Geology) CAREER Carnegie Museum of Natural History (CMNH) Laboratory Technician, Section of Vertebrate Paleontology, 1979 Research Assistant, Section of Vertebrate Paleontology, 1980 Curatorial Assistant, Section of Vertebrate Paleontology, 1980-1984 Scientific Preparator, Section of Paleobotany, 1985-1986 Scientific Preparator, Section of Vertebrate Paleontology, 1985-2002 Acting Collection Manager/Scientific Preparator, 2003-2004 Collection Manager, 2005-present PALEONTOLOGICAL FIELD EXPERIENCE Late Pennsylvanian through Early Permian of Colorado, New Mexico and Utah (fish, amphibians and reptiles) Early Permian of Germany, Bromacker quarry (amphibians and reptiles) Triassic of New Mexico, Coelophysis quarry (Coelophysis and other reptiles) Upper Jurassic of Colorado (mammals and herps) Tertiary of Montana, Nevada, and Wyoming (mammals and herps) Pleistocene of West Virginia (mammals and herps) Lake sediment cores and lake sediment surface samples, Wyoming (pollen and seeds) PROFESSIONAL APPOINTMENTS Associate Editor, Society of Vertebrate Paleontology, 1998-2000. Research Associate in the Science Division, New Mexico Museum of Natural History and Science, 2007-present. PROFESSIONAL ASSOCIATIONS Society of Vertebrate Paleontology Paleontological Society LECTURES and TUTORIALS (Invited and public) 1994. Middle Eocene frogs from central Wyoming: ontogeny and taphonomy. California State University, San Bernardino 1994. Mechanical preparation of vertebrate fossils. California State University, San Bernardino 1994. Mechanical preparation of vertebrate fossils. University of Chicago 2001. -

OP-MOLB160269 Online 744..771
Evolutionary History of the Asian Horned Frogs (Megophryinae): Integrative Approaches to Timetree Dating in the Absence of a Fossil Record Stephen Mahony,*,1,2 Nicole M. Foley,1 S.D. Biju,2 and Emma C. Teeling*,1 1School of Biology and Environmental Science, University College Dublin, Belfield, Dublin, Ireland 2Systematics Lab, Department of Environmental Studies, University of Delhi, Delhi, India *Corresponding authors: E-mails: [email protected]; [email protected]. Associate editor: Beth Shapiro Abstract Downloaded from https://academic.oup.com/mbe/article-abstract/34/3/744/2919384 by guest on 06 August 2019 Molecular dating studies typically need fossils to calibrate the analyses. Unfortunately, the fossil record is extremely poor or presently nonexistent for many species groups, rendering such dating analysis difficult. One such group is the Asian horned frogs (Megophryinae). Sampling all generic nomina, we combined a novel 5 kb dataset composed of four nuclear and three mitochondrial gene fragments to produce a robust phylogeny, with an extensive external morpho- logical study to produce a working taxonomy for the group. Expanding the molecular dataset to include out-groups of fossil-represented ancestral anuran families, we compared the priorless RelTime dating method with the widely used prior-based Bayesian timetree method, MCMCtree, utilizing a novel combination of fossil priors for anuran phylogenetic dating. The phylogeny was then subjected to ancestral phylogeographic analyses, and dating estimates were compared with likely biogeographic vicariant events. Phylogenetic analyses demonstrated that previously proposed systematic hypotheses were incorrect due to the paraphyly of genera. Molecular phylogenetic, morphological, and timetree results support the recognition of Megophryinae as a single genus, Megophrys, with a subgenus level classification. -

Exceptional Fossil Preservation During CO2 Greenhouse Crises? Gregory J
Palaeogeography, Palaeoclimatology, Palaeoecology 307 (2011) 59–74 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Exceptional fossil preservation during CO2 greenhouse crises? Gregory J. Retallack Department of Geological Sciences, University of Oregon, Eugene, Oregon 97403, USA article info abstract Article history: Exceptional fossil preservation may require not only exceptional places, but exceptional times, as demonstrated Received 27 October 2010 here by two distinct types of analysis. First, irregular stratigraphic spacing of horizons yielding articulated Triassic Received in revised form 19 April 2011 fishes and Cambrian trilobites is highly correlated in sequences in different parts of the world, as if there were Accepted 21 April 2011 short temporal intervals of exceptional preservation globally. Second, compilations of ages of well-dated fossil Available online 30 April 2011 localities show spikes of abundance which coincide with stage boundaries, mass extinctions, oceanic anoxic events, carbon isotope anomalies, spikes of high atmospheric carbon dioxide, and transient warm-wet Keywords: Lagerstatten paleoclimates. Exceptional fossil preservation may have been promoted during unusual times, comparable with fi Fossil preservation the present: CO2 greenhouse crises of expanding marine dead zones, oceanic acidi cation, coral bleaching, Trilobite wetland eutrophication, sea level rise, ice-cap melting, and biotic invasions. Fish © 2011 Elsevier B.V. All rights reserved. Carbon dioxide Greenhouse 1. Introduction Zeigler, 1992), sperm (Nishida et al., 2003), nuclei (Gould, 1971)and starch granules (Baxter, 1964). Taphonomic studies of such fossils have Commercial fossil collectors continue to produce beautifully pre- emphasized special places where fossils are exceptionally preserved pared, fully articulated, complex fossils of scientific(Simmons et al., (Martin, 1999; Bottjer et al., 2002). -
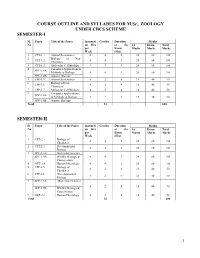
COURSE OUTLINE and SYLLABUS for M.Sc., ZOOLOGY UNDER CBCS SCHEME SEMESTER-I
COURSE OUTLINE AND SYLLABUS FOR M.Sc., ZOOLOGY UNDER CBCS SCHEME SEMESTER-I Sl. Paper Title of the Paper Instructi Credits Duration Marks No on Hrs of the IA Exam Total per Exam Marks Marks Marks Week (Hrs) 1 CPT-1.1 Animal Systematics 4 4 3 20 80 100 2 Biology of Non- CPT-1.2 4 4 3 20 80 100 Chordates 3 CPT-1.3 Molecular Cell Biology 4 4 3 20 80 100 4 Computer Applications & SPT-1.4A Methods in Biology 4 4 3 20 80 100 SPT-1.4B Aquatic Biology 5 CPP-1.5 Animal Systematics 4 2 4 10 40 50 6 Biology of Non- CPP-1.6 4 2 4 10 40 50 Chordates 7 CPP-1.7 Molecular Cell Biology 4 2 4 10 40 50 8 Computer Applications SPP-1.8A & Methods in Biology 4 2 4 10 40 50 SPP-1.8B Aquatic Biology Total 24 600 SEMESTER-II Sl. Paper Title of the Paper Instructi Credits Duration Marks No on Hrs of the IA Exam Total per Exam Marks Marks Marks Week (Hrs) 1 CPT-2.1 Biology of 4 4 3 20 80 100 Chordates 2 CPT-2.2 Developmental 4 4 3 20 80 100 Biology 3 SPT-2.3A Molecular Genetics SPT-2.3B Wildlife Biology & 4 4 3 20 80 100 Conservation 4 OET-2.4 Human Physiology 4 4 3 20 80 100 5 CPP-2.5 Biology of 4 2 4 10 40 50 Chordates 6 CPP-2.6 Developmental 4 2 4 10 40 50 Biology 7 SPP-2.7A Molecular Genetics 4 2 4 10 40 50 SPP-2.7B Wildlife Biology & Conservation 8 OEP-2.8 Human Physiology 4 2 4 10 40 50 Total 24 600 1 SEMESTER-III Sl. -

Larval Development in Oligocene Palaeobatrachid Frogs
Larval development in Oligocene palaeobatrachid frogs ZBYNĚK ROČEK Roček, Z. 2003. Larval development in Oligocene palaeobatrachid frogs. Acta Palaeontologica Polonica 48 (4): 595–607. A detailed account of the development of skeletal and some soft−tissue structures is based on 171 fossil tadpoles and meta− morphosing froglets of Palaeobatrachus sp. from the Late Oligocene of the Czech Republic (locality Bechlejovice). Their exceptionally good preservation resulted from fossilization in diatomites. The fossil developmental series was com− pared with normal development of the contemporary anuran Xenopus laevis (Pipidae) represented by cleared and stained (alizarin/toluidin−blue) whole−mount specimens. The comparison revealed that in spite of differences in the sequence of ossification and its timing (e.g., ossification of the otic capsules and ribs was retarded in Xenopus whereas dermal ossifi− cation was retarded in Palaeobatrachus), in the number of free ribs, and in composition of the sacral region (the synsacrum in Palaeobatrachus involves two posterior presacrals, whereas there is a single sacral in Xenopus), both gen− era were similar in great number of anatomical features that appear during development. The most important difference is the shape of vertebral centrum (procoelous in Palaeobatrachus, opisthocoelous in all Pipidae) which is formed in com− paratively early developmental stages. A view that could result from anatomical comparisons is that Palaeobatrachus could be derived from the Pipidae, but this is doubtful due to biostratigraphic and palaeogeographic discrepancies. The earliest palaeobatrachids were recorded from the Late Cretaceous of Europe but pipids could not invade northern conti− nents after the Early Cretaceous when the Tethys Sea prevented interchanges of anuran faunas. -

Frontoparietal Bone in Extinct Palaeobatrachidae (Anura): Its Variation and Taxonomic Value
THE ANATOMICAL RECORD 298:1848–1863 (2015) Frontoparietal Bone in Extinct Palaeobatrachidae (Anura): Its Variation and Taxonomic Value ZBYNEK ROCEK, 1* RENAUD BOISTEL,2 NICOLAS LENOIR,3 ARNAUD MAZURIER,4 STEPHANIE E. PIERCE,5 JEAN-CLAUDE RAGE,6 SERGEI V. SMIRNOV,7 ACHIM H. SCHWERMANN,8 XAVIER VALENTIN,2 9 10 11 MARTON VENCZEL, MICHAEL WUTTKE, AND TOMA S ZIKMUND 1Department of Palaeobiology, Geological Institute, Academy of Sciences of the Czech Republic, Prague, Czech Republic 2Institut International de Paleoprimatologie et de Paleontologie Humaine, UMR 7262 CNRS, Universite de Poitiers, Poitiers, France 3Multiscale Group, Laboratoire Navier, UMR8205-CNRS/ENPC/IFSTTAR/Universite Paris-Est, Champs-sur-Marne, France 4Institut de Chimie des Milieux et Materiaux de Poitiers, UMR 7285 Universitede Poitiers, UFR SFA, Poitiers, France 5Department of Organismic and Evolutionary Biology and Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts 6Sorbonne Universites—CR2P—MNHN, CNRS, UPMC-Paris 6, Museum National d’Histoire Naturelle, Paris, France 7Laboratory of Evolutionary Morphology, a.N. Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, Moscow, Russia 8Steinmann-Institut fur€ Geologie, Mineralogie und Palaontologie,€ Universitat€ Bonn, Bonn, Germany 9 T; arii Cris¸urilor Museum, Oradea, Romania 10Department of Archaeology, General Department of Cultural Heritage Rhineland Palatinate, Section Geological History of the Earth, Mainz, Germany 11X-Ray Micro CT and Nano CT Research Group, CEITEC—Central European Institute of Technology, Brno University of Technology, Brno, Czech Republic ABSTRACT Palaeobatrachidae are extinct frogs from Europe closely related to the Gondwanan Pipidae, which includes Xenopus. Their frontoparietal is a distinctive skeletal element which has served as a basis for establishing the genus Albionbatrachus. -

Neogene Amphibians and Reptiles (Caudata, Anura, Gekkota, Lacertilia, and Testudines) from the South of Western Siberia, Russia, and Northeastern Kazakhstan
Neogene amphibians and reptiles (Caudata, Anura, Gekkota, Lacertilia, and Testudines) from the south of Western Siberia, Russia, and Northeastern Kazakhstan Davit Vasilyan1,2,3, Vladimir S. Zazhigin4 and Madelaine Bo¨hme1,5 1 Department of Geosciences, Eberhard Karls University Tu¨bingen, Tu¨bingen, Germany 2 JURASSICA Museum, Porrentruy, Switzerland 3 Department of Geosciences, University of Fribourg, Fribourg, Switzerland 4 Institute of Geology, Russian Academy of Sciences, Moscow, Russia 5 Senckenberg Center for Human Evolution and Palaeoecology, Eberhard Karls University Tu¨bingen, Tu¨bingen, Germany ABSTRACT Background: The present-day amphibian and reptile fauna of Western Siberia are the least diverse of the Palaearctic Realm, as a consequence of the unfavourable climatic conditions that predominate in this region. The origin and emergence of these herpetofaunal groups are poorly understood. Aside from the better-explored European Neogene localities yielding amphibian and reptile fossil remains, the Neogene herpetofauna of Western Asia is understudied. The few available data need critical reviews and new interpretations, taking into account the more recent records of the European herpetofauna. The comparison of this previous data with that of European fossil records would provide data on palaeobiogeographic affiliations of the region as well as on the origin and emergence of the present-day fauna of Western Siberia. An overview of the earliest occurrences of certain amphibian lineages is still needed. In addition, studies that address such knowledge gaps can be Submitted 16 August 2016 useful for molecular biologists in their calibration of molecular clocks. Accepted 24 January 2017 Methods and Results: In this study, we considered critically reviewed available data Published 23 March 2017 from amphibian and reptile fauna from over 40 Western Siberian, Russian and Corresponding author Northeastern Kazakhstan localities, ranging from the Middle Miocene to Early Davit Vasilyan, [email protected] Pleistocene. -

Retallack 2011 Lagerstatten
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright Author's personal copy Palaeogeography, Palaeoclimatology, Palaeoecology 307 (2011) 59–74 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Exceptional fossil preservation during CO2 greenhouse crises? Gregory J. Retallack Department of Geological Sciences, University of Oregon, Eugene, Oregon 97403, USA article info abstract Article history: Exceptional fossil preservation may require not only exceptional places, but exceptional times, as demonstrated Received 27 October 2010 here by two distinct types of analysis. First, irregular stratigraphic spacing of horizons yielding articulated Triassic Received in revised form 19 April 2011 fishes and Cambrian trilobites is highly correlated in sequences in different parts of the world, as if there were Accepted 21 April 2011 short temporal intervals of exceptional preservation globally. Second, compilations of ages of well-dated fossil Available online 30 April 2011 localities show spikes of abundance which coincide with stage boundaries, mass extinctions, oceanic anoxic events, carbon isotope anomalies, spikes of high atmospheric carbon dioxide, and transient warm-wet Keywords: Lagerstatten paleoclimates.