Prickly Ash Seed Kernel: a New Bio-Fumigation Material Against Tobacco Black Shank
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A New Indicator to Assess Public Perception of Air Pollution Based on Complaint Data
applied sciences Article A New Indicator to Assess Public Perception of Air Pollution Based on Complaint Data Yong Sun 1 , Fengxiang Jin 2, Yan Zheng 1, Min Ji 1,* and Huimeng Wang 2,3 1 College of Geodesy and Geomatics, Shandong University of Science and Technology, Qingdao 266590, China; [email protected] (Y.S.); [email protected] (Y.Z.) 2 College of Surveying and Geo-Informatics, Shandong Jianzhu University, Ji’nan 250101, China; [email protected] (F.J.); [email protected] (H.W.) 3 State Key Laboratory of Resources and Environmental Information System, Institute of Geographic Science and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China * Correspondence: [email protected] Abstract: Severe air pollution problems have led to a rise in the Chinese public’s concern, and it is necessary to use monitoring stations to monitor and evaluate pollutant levels. However, monitoring stations are limited, and the public is everywhere. It is also essential to understand the public’s awareness and behavioral response to air pollution. Air pollution complaint data can more directly reflect the public’s real air quality perception than social media data. Therefore, based on air pollution complaint data and sentiment analysis, we proposed a new air pollution perception index (APPI) in this paper. Firstly, we constructed the emotional dictionary for air pollution and used sentiment analysis to calculate public complaints’ emotional intensity. Secondly, we used the piecewise function to obtain the APPI based on the complaint Kernel density and complaint emotion Kriging interpolation, and we further analyzed the change of center of gravity of the APPI. -

Proposed Re-Election and Appointment of Directors and Supervisors
Hong Kong Exchanges and Clearing Limited and The Stock Exchange of Hong Kong Limited take no responsibility for the contents of this announcement, make no representation as to its accuracy or completeness and expressly disclaim any liability whatsoever for any loss howsoever arising from or in reliance upon the whole or any part of the contents of this announcement. LUZHENG FUTURES Company Limited 魯証期貨股份有限公司 (A joint stock company incorporated in the People’s Republic of China with limited liability) (Stock Code: 01461) PROPOSED RE-ELECTION AND APPOINTMENT OF DIRECTORS AND SUPERVISORS PROPOSED RE-ELECTION AND APPOINTMENT OF DIRECTORS According to the requirements of the Company Law of the People’s Republic of China and the Articles of Association (the “Articles of Association”) of LUZHENG FUTURES Company Limited (the “Company”), the term of office of directors of the Company (the “Director(s)”) for each session is three years. Upon the expiration of the term of office, a Director shall be eligible for re-election and re-appointment. Directors who are not staff representatives shall be elected and removed by shareholders at general meetings, while Directors as staff representatives shall be elected and removed through democratic means by the staff of the Company, details of which will be announced separately. On 24 October 2019, the board of Directors of the Company (the “Board”) passed the resolutions for the proposed re-election and appointment of non-staff representative members of the third session of the Board which shall be subject to the approval at the forthcoming first extraordinary general meeting of the Company for the year 2019 (the “EGM”). -

Cereal Series/Protein Series Jiangxi Cowin Food Co., Ltd. Huangjindui
产品总称 委托方名称(英) 申请地址(英) Huangjindui Industrial Park, Shanggao County, Yichun City, Jiangxi Province, Cereal Series/Protein Series Jiangxi Cowin Food Co., Ltd. China Folic acid/D-calcium Pantothenate/Thiamine Mononitrate/Thiamine East of Huangdian Village (West of Tongxingfengan), Kenli Town, Kenli County, Hydrochloride/Riboflavin/Beta Alanine/Pyridoxine Xinfa Pharmaceutical Co., Ltd. Dongying City, Shandong Province, 257500, China Hydrochloride/Sucralose/Dexpanthenol LMZ Herbal Toothpaste Liuzhou LMZ Co.,Ltd. No.282 Donghuan Road,Liuzhou City,Guangxi,China Flavor/Seasoning Hubei Handyware Food Biotech Co.,Ltd. 6 Dongdi Road, Xiantao City, Hubei Province, China SODIUM CARBOXYMETHYL CELLULOSE(CMC) ANQIU EAGLE CELLULOSE CO., LTD Xinbingmaying Village, Linghe Town, Anqiu City, Weifang City, Shandong Province No. 569, Yingerle Road, Economic Development Zone, Qingyun County, Dezhou, biscuit Shandong Yingerle Hwa Tai Food Industry Co., Ltd Shandong, China (Mainland) Maltose, Malt Extract, Dry Malt Extract, Barley Extract Guangzhou Heliyuan Foodstuff Co.,LTD Mache Village, Shitan Town, Zengcheng, Guangzhou,Guangdong,China No.3, Xinxing Road, Wuqing Development Area, Tianjin Hi-tech Industrial Park, Non-Dairy Whip Topping\PREMIX Rich Bakery Products(Tianjin)Co.,Ltd. Tianjin, China. Edible oils and fats / Filling of foods/Milk Beverages TIANJIN YOSHIYOSHI FOOD CO., LTD. No. 52 Bohai Road, TEDA, Tianjin, China Solid beverage/Milk tea mate(Non dairy creamer)/Flavored 2nd phase of Diqiuhuanpo, Economic Development Zone, Deqing County, Huzhou Zhejiang Qiyiniao Biological Technology Co., Ltd. concentrated beverage/ Fruit jam/Bubble jam City, Zhejiang Province, P.R. China Solid beverage/Flavored concentrated beverage/Concentrated juice/ Hangzhou Jiahe Food Co.,Ltd No.5 Yaojia Road Gouzhuang Liangzhu Street Yuhang District Hangzhou Fruit Jam Production of Hydrolyzed Vegetable Protein Powder/Caramel Color/Red Fermented Rice Powder/Monascus Red Color/Monascus Yellow Shandong Zhonghui Biotechnology Co., Ltd. -

Table of Codes for Each Court of Each Level
Table of Codes for Each Court of Each Level Corresponding Type Chinese Court Region Court Name Administrative Name Code Code Area Supreme People’s Court 最高人民法院 最高法 Higher People's Court of 北京市高级人民 Beijing 京 110000 1 Beijing Municipality 法院 Municipality No. 1 Intermediate People's 北京市第一中级 京 01 2 Court of Beijing Municipality 人民法院 Shijingshan Shijingshan District People’s 北京市石景山区 京 0107 110107 District of Beijing 1 Court of Beijing Municipality 人民法院 Municipality Haidian District of Haidian District People’s 北京市海淀区人 京 0108 110108 Beijing 1 Court of Beijing Municipality 民法院 Municipality Mentougou Mentougou District People’s 北京市门头沟区 京 0109 110109 District of Beijing 1 Court of Beijing Municipality 人民法院 Municipality Changping Changping District People’s 北京市昌平区人 京 0114 110114 District of Beijing 1 Court of Beijing Municipality 民法院 Municipality Yanqing County People’s 延庆县人民法院 京 0229 110229 Yanqing County 1 Court No. 2 Intermediate People's 北京市第二中级 京 02 2 Court of Beijing Municipality 人民法院 Dongcheng Dongcheng District People’s 北京市东城区人 京 0101 110101 District of Beijing 1 Court of Beijing Municipality 民法院 Municipality Xicheng District Xicheng District People’s 北京市西城区人 京 0102 110102 of Beijing 1 Court of Beijing Municipality 民法院 Municipality Fengtai District of Fengtai District People’s 北京市丰台区人 京 0106 110106 Beijing 1 Court of Beijing Municipality 民法院 Municipality 1 Fangshan District Fangshan District People’s 北京市房山区人 京 0111 110111 of Beijing 1 Court of Beijing Municipality 民法院 Municipality Daxing District of Daxing District People’s 北京市大兴区人 京 0115 -

December 1998
JANUARY - DECEMBER 1998 SOURCE OF REPORT DATE PLACE NAME ALLEGED DS EX 2y OTHER INFORMATION CRIME Hubei Daily (?) 16/02/98 04/01/98 Xiangfan C Si Liyong (34 yrs) E 1 Sentenced to death by the Xiangfan City Hubei P Intermediate People’s Court for the embezzlement of 1,700,00 Yuan (US$20,481,9). Yunnan Police news 06/01/98 Chongqing M Zhang Weijin M 1 1 Sentenced by Chongqing No. 1 Intermediate 31/03/98 People’s Court. It was reported that Zhang Sichuan Legal News Weijin murdered his wife’s lover and one of 08/05/98 the lover’s relatives. Shenzhen Legal Daily 07/01/98 Taizhou C Zhang Yu (25 yrs, teacher) M 1 Zhang Yu was convicted of the murder of his 01/01/99 Zhejiang P girlfriend by the Taizhou City Intermediate People’s Court. It was reported that he had planned to kill both himself and his girlfriend but that the police had intervened before he could kill himself. Law Periodical 19/03/98 07/01/98 Harbin C Jing Anyi (52 yrs, retired F 1 He was reported to have defrauded some 2600 Liaoshen Evening News or 08/01/98 Heilongjiang P teacher) people out of 39 million Yuan 16/03/98 (US$4,698,795), in that he loaned money at Police Weekend News high rates of interest (20%-60% per annum). 09/07/98 Southern Daily 09/01/98 08/01/98 Puning C Shen Guangyu D, G 1 1 Convicted of the murder of three children - Guangdong P Lin Leshan (f) M 1 1 reported to have put rat poison in sugar and 8 unnamed Us 8 8 oatmeal and fed it to the three children of a man with whom she had a property dispute. -

The First International U3as Online Art Awards 2020 ---Creativity Winners
The First International U3As Online Art Awards 2020 ---Creativity Winners list / Premier Concours International d'art des U3As 2020 --- Liste des gagnants en créativité Nationality/N Awards/ Prix Participants U3A ationalité Top Awards/ Meilleur prix Noel Bird Noosa Queensland Australian Golden Awards/ Prix or Yu Desong Yantai Tianma Vellas U3A Chinese Xie Yong Sishui County U3A Chinese Silver Awards/ Prix argent Daniela Lestinska UTA EUBA Bratislava Slovakia Slovikian Nibale Salloum U3A AUT Lebanon Lebanese BEST Carving Awards/ Lu Yingxuan Qingdao West Coast New District U3A Chinese Prix sculpture Cao Changxin Qingdao West Coast New District U3A Chinese Excellence Awards/ Prix excellence Zhang Linyan Yantai Tianma Vellas U3A Chinese Wang Weiting Yantai Zhaoyuan U3A Chinese BEST Carving Awards/ Prix sculpture Excellence Awards/ Prix excellence Li Huayue Yantai Zhaoyuan U3A Chinese Golden Awards/ Prix or Wang Xiangqin Qingdao Municipal U3A Chinese Zhang Jinping Yantai Municipal U3A Chinese Silver Awards/ Prix argent N.Narantuya Long Life Academy Mongolian Tian Shoufeng Zibo Yiyuan County U3A Chinese BEST Color Awards/ Qiu Yushui Tai'an Municipal U3A Chinese Prix couleur Wang Jun Jinan University for Seniors Chinese Excellence Awards/ Prix excellence Zhuang Xiuhua Qingdao Municipal U3A Chinese Guo Huanhua Qingdao Municipal U3A Chinese Wang Guojun Yantai Municipal U3A Chinese Golden Awards/ Prix or Qin Xiaoding Rudong County U3A, Jiangsu Province Chinese Zhang Yanan Yantai Tianma Vellas U3A Chinese Silver Awards/ Prix argent BEST Expression -

Analysis of the Influence of Rural Land Stock Cooperative System on Farmers' Income —A Case Study of Farmers in Zaozhuang City
Advances in Social Science, Education and Humanities Research (ASSEHR), volume 65 2016 International Conference on Education, Management Science and Economics (ICEMSE-16) Analysis of the Influence of Rural Land Stock Cooperative System on Farmers' Income —A Case Study of Farmers in Zaozhuang City Zhang Lin College of Economics and Management Zaozhuang University, Zaozhuang 277100, China College of Economics and Management China Agricultural University Beijing, 100083, China [email protected] Abstract—Basing on the field investigation of the Zaozhuang evaluate the farmer's income effect of the rural land stock city in Shandong Province, this paper made an empirical study cooperative system. It cannot be denied that after the on the impact of rural land share cooperative system on the implementation of rural land joint-stock cooperative system, income of peasant households. The conclusion is that the rural many farmers' income levels are significantly higher than land stock cooperative can increase peasants' total income, non- before, [3]But whether this increase is statistically significantly? agricultural income per capita. The paper also analyzed the To what degree that the farmers’ income increases result from problems caused in the operation of rural land stock cooperative the effects of the rural land stock cooperative system? In view system. At the end of the paper, the solution with specific aim is of this, based on the field survey of farmers in Zaozhuang city proposed. of Shandong province, this article expand empirical study on Keywords—the Land Stock Cooperative System, Influence, the income effect of the rural land stock cooperative system. Income, Problems, Zaozhuang City II. -

Resettlement Planning Document PRC: Risk Mitigation And
Resettlement Planning Document Document Stage: Draft for Consultation Project Number: 40683 March 2010 PRC: Risk Mitigation and Strengthening of Endangered Reservoirs in Shandong Province Project Due Diligence Report for Shizuizi Reservoir Subproject in Zaozhuang City (English) Prepared by the Shandong Provincial Government. The resettlement plan is a document of the borrower. The views expressed herein do not necessarily represent those of ADB’s Board of Directors, Management, or staff, and may be preliminary in nature. Zaozhuang City Shizuizi Reservoir Risk Mitigation Project Under Risk Mitigation of Endangered Reservoir Project in Shandong Province Of The People’s Republic of China Due Diligence Report Design Institute of Zaozhuang City Water Resources Bureau of Shandong Province 11 November 2009 Letter of Endorsement Zaozhuang City Water Resources Bureau (ZCWRb) received the approval of constructing the Risk Mitigation of Shizuizi Reservoir Project in Zaozhuang City from the related departments. This Project is proposed to be started in November 2009, and completed by November 2010. Zaohuang City Government, through Ministry of Finance, has requested a loan from the Asian Development Bank (ADB) to finance part of the Project. Accordingly, the Project will be implemented in compliance with ADB social safeguard policies. This Due Diligence Report represents a key requirement of ADB and will constitute the basis for land acquisition and resettlement. The due diligence report fully complies with requirements of the relevant laws, regulations and policies of the People’s Republic of China (PRC), Shandong Province and Zaozhuang City Government as well as complies with ADB’s policy on involuntary resettlement. Zaozhuang City Government and ZCWRb hereby affirm the contents of this Due Diligence Report prepared dated in November 2009 and ensures that the resettlement will be made available as stipulated in the budget.ZCWRb has discussed the draft resettlement Plan with relevant local officials and has obtained their concurrence. -
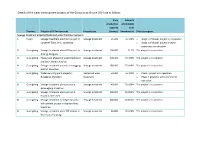
Details of the Water Development Projects of the Group As at 30 June 2014 Are As Follows
Details of the water development projects of the Group as at 30 June 2014 are as follows: Daily Interests production attributable capacity to SI Province Projects of SI Environment Project type (tonnes) Environment Project progress Sewage treatment projects/Reclaimed water treatment projects 1 Fujian Sewage treatment plant BOT project in Sewage treatment 25,000 76.419% Stage 1 of phase I project is in operation. Longmen Town, Anxi, Quanzhou Stage 2 of phase I project is yet to commence construction. 2 Guangdong Sewage treatment plant BOT project in Sewage treatment 100,000 75.5% The project is in operation. Dalang, Donguan 3 Guangdong Phase I and phase II of water treatment Sewage treatment 200,000 76.419% The project is in operation. project in Meihu, Huizhou 4 Guangdong Sewage treatment projects in Longgang Sewage treatment 280,000 76.419% The project is in operation. District, Shenzhen 5 Guangdong Water recycling plant project in Reclaimed water 50,000 76.419% Phase I project is in operation. Henggang, Shenzhen treatment Phase II project is yet to commence operation. 6 Guangdong Sewage treatment plant project in Sewage treatment 40,000 76.419% The project is in operation. Banxuegang, Shenzhen 7 Guangdong Sewage treatment plant project in Sewage treatment 260,000 45.851% The project is in operation. Guanlan, Shenzhen 8 Guangdong Sewage treatment contingency works Sewage treatment 400,000 45.851% The project is in operation. entrustment project in Guanlan River, Shenzhen 9 Guangdong Sewage treatment plant TOT project in Sewage treatment 40,000 76.419% The project is in operation. Wuchuan, Zhanjiang Daily Interests production attributable Projects of SI Environment capacity to SI Province (continued) Project type (tonnes) Environment Project progress 10 Guangxi City sewage treatment plant BOT Sewage treatmen 40,000 75.5% The project is in operation. -
Bestseller Factory List
BESTSELLER FACTORY LIST LAST UPDATED: 21.05.2021 It is imperative for us to work with our partners in an open and honest way. We continuously seek to create more transparency in our supply chain to address risks and promote positive change. In light of this, and to provide increased transparency, we are making supplier factory information publicly available. The factory list includes the name, address, product type and number of workers of all tier 1 manufacturing factories (cut-make-trim) of apparel, footwear and accessories. The list will be updated twice a year. As changes occur in our supply chain with new partners, new factories onboarded, or factories being phased out, such alterations will be collected and published in the next supplier factory list update. FACTORY NAME ADDRESS MALE FEMALE NO. OF POSTAL CITY REGION COUNTRY PRODUCT EMPLOYEE EMPLOYEE EMPLOYEES CODE TYPES % % Almeg Shoes No. 14 Rruga Albania 5 95 <500 2000 Durres Durres ALBANIA Footwear Aba Fashions Ltd. No. 521/1 Gacha, Gazipur City Corporation 35 65 1001-5000 1704 Gazipur Dhaka BANGLADESH Apparel ABM Fashions Ltd. No.1143 &1145 Kashimpur Road, Konabari, Gazipur 30 70 1001-5000 1700 Dhaka Dhaka BANGLADESH Apparel Agami Apparels Ltd. Nayapara, Kathgara, Ashulia 40 60 5001-10000 1344 Savar Dhaka BANGLADESH Apparel Agami Fashions Ltd. Abdus Sattar Road, Pallibiduyt, Holding No. 79/1 30 70 1001-5000 1751 Gazipur Dhaka BANGLADESH Apparel AKM Knit Wear Ltd. No. 14 Gedda Karnapara, Ulail, Savar 35 65 >10000 1340 Dhaka Dhaka BANGLADESH Apparel Al-Muslim Apparels Ltd. No. 12 Nischintapur 25 75 <500 1341 Ashulia Dhaka BANGLADESH Apparel Aman Tex Ltd. -

Original Article the Effect of Mirna-210 on the Biological Behavior Ovarian Carcinoma A2780 Cells
Int J Clin Exp Med 2017;10(1):498-506 www.ijcem.com /ISSN:1940-5901/IJCEM0039350 Original Article The effect of miRNA-210 on the biological behavior ovarian carcinoma A2780 cells Yang Zhang1, Yan Sun2, Lin Lin3, Chenchen Ding1 Departments of 1Gynecology and Obstetrics, 3Neonatology, Tengzhou Central People’s Hospital, Tengzhou 277599, P. R. China; 2Department of Gynecology and Obstetrics, Shanting District People’s Hospital, Zaozhuang 277200, P. R. China Received September 3, 2016; Accepted September 19, 2016; Epub January 15, 2017; Published January 30, 2017 Abstract: Objective: To explore the expression of miRNA-210 (miR-210) in human ovarian cancer and the effect of silencing of miR-210 on the biological behavior of A2780 cells. Methods: Tissue specimens of 20 patients suffer- ing from ovarian cancer hospitalized in our hospital from June 2010 to June 2012 were collected. Real-time PCR was used to detect the expression of miR-210 in ovarian cancer and para-cancer tissues, as well as ovarian cancer cell line A2780 and normal ovarian cell line IOSE80. After miR-210 inhibitor was transfected into A2780 cells by LipofectamineTM 2000, the proliferation of A2780 cells was detected by MTT and soft agar colony formation assay, cell cycle and apoptosis by flow cytometry and cells’ migration and invasiveness by Transwell assay. A tumor model was established by inoculating A2780, A2780/NC and A2780/210 cells into BALB/C nude mice subcutaneously. Results: The expression level of miR-210 in ovarian cancer tissues and A2780 cells was evidently higher than that in para-cancer tissues and normal ovary cells (P<0.01). -

CHINA the Church of Almighty God: Prisoners Database (1663 Cases)
CHINA The Church of Almighty God: Prisoners Database (1663 cases) Prison term: 15 years HE Zhexun Date of birth: On 18th September 1963 Date and place of arrest: On 10th March 2009, in Xuchang City, Henan Province Charges: Disturbing social order and using a Xie Jiao organization to undermine law enforcement because of being an upper-level leader of The Church of Almighty God in mainland China, who was responsible for the overall work of the church Statement of the defendant: He disagreed with the decision and said what he believed in is not a Xie Jiao. Court decision: In February 2010, he was sentenced to 15 years in prison by the Zhongyuan District People’s Court of Zhengzhou City, Henan Province. Place of imprisonment: No. 1 Prison of Henan Province Other information: He was regarded by the Chinese authorities as a major criminal of the state and had long been on the wanted list. To arrest him, authorities offered 500,000 RMB as a reward to informers who gave tips leading to his arrest to police. He was arrested at the home of a Christian in Xuchang City, Henan Province. Based on the information from a Christian serving his sentence in the same prison, HE Zhexun was imprisoned in a separate area and not allowed to contact other prisoners. XIE Gao, ZOU Yuxiong, SONG Xinling and GAO Qinlin were arrested in succession alongside him and sentenced to prison terms ranging from 11 to 12 years. Source: https://goo.gl/aGkHBj Prison term: 14 years MENG Xiumei Age: Forty-one years old Date and place of arrest: On 14th August 2014, in Xinjiang Uyghur Autonomous Region Charges: Using a Xie Jiao organization to undermine law enforcement because of being a leader of The Church of Almighty God and organizing gatherings for Christians and the work of preaching the gospel in Ili prefecture Statement of the defendant: She claimed that her act did not constitute crimes.