Inferring the Social Organization of Medieval Upper Nubia Using Nonmetric Traits of the Skull
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Research on Ancient DNA in the Near East Mateusz Baca*1, Martyna Molak2 1 Center for Precolumbian Studies, University of Warsaw, Ul
Bioarchaeology of the Near East, 2:39–61 (2008) Research on ancient DNA in the Near East Mateusz Baca*1, Martyna Molak2 1 Center for Precolumbian Studies, University of Warsaw, ul. Krakowskie Przedmieście 26/28, 00-927 Warsaw, Poland email: [email protected] (corresponding author) 2 Institute of Genetics and Biotechnology, University of Warsaw, ul. Pawińskiego 5a, 05-106 Warsaw, Poland Abstract: In the early 1990s, when studies of ancient DNA became possible, new perspectives of analyzing archaeological data also developed. Nowadays, because the methodology related to ancient DNA research is well developed, it has been used to reveal several aspects of human history and interaction. Here we review the basic concepts, methodologies, and recent developments in the fi eld of ancient DNA studies with a special refe- rence to the Near East. Th is includes not only human but also animal and bacterial DNA. Key words: archaeogenetics, aDNA, mtDNA, tuberculosis, animal domestication Introduction Human genomes accumulate mutations gradually over time. Th e forces of genetic drift and natural selection either cause these changes to disappear or to become established in the popu- lation. By the end of the 1990s, Amorim (1999) introduced the term “archaeogenetics” in reference to using information regarding genetic diff erences between humans to understand demographic events that took place in the past. Pioneering studies of human genetic diversity date back to 1970s when Cavalli-Sforza published a report on the diversity of European populations based on classic protein mark- ers (see Cavalli-Sforza et al. 1994 for a review). In the mid-eighties, great opportunities for studying human diversity arose with the invention of polymerase chain reaction (PCR). -

African Journal of History and Culture
OPEN ACCESS African Journal of History and Culture March 2019 ISSN: 2141-6672 DOI: 10.5897/AJHC www.academicjournals.org Editors Pedro A. Fuertes-Olivera Ndlovu Sabelo University of Valladolid Ferguson Centre for African and Asian Studies, E.U.E. Empresariales ABOUTOpen University, AJHC Milton Keynes, Paseo del Prado de la Magdalena s/n United Kingdom. 47005 Valladolid Spain. Biodun J. Ogundayo, PH.D The African Journal of History and Culture (AJHC) is published monthly (one volume per year) by University of Pittsburgh at Bradford Academic Journals. Brenda F. McGadney, Ph.D. 300 Campus Drive School of Social Work, Bradford, Pa 16701 University of Windsor, USA. Canada. African Journal of History and Culture (AJHC) is an open access journal that provides rapid publication Julius O. Adekunle (monthly) of articles in all areas of the subject. TheRonen Journal A. Cohenwelcomes Ph.D. the submission of manuscripts Department of History and Anthropology that meet the general criteria of significance andDepartment scientific of excellence.Middle Eastern Papers and will be published Monmouth University Israel Studies / Political Science, shortlyWest Long after Branch, acceptance. NJ 07764 All articles published in AJHC are peer-reviewed. Ariel University Center, USA. Ariel, 40700, Percyslage Chigora Israel. Department Chair and Lecturer Dept of History and Development Studies Midlands State University ContactZimbabwe Us Private Bag 9055, Gweru, Zimbabwe. Editorial Office: [email protected] Help Desk: [email protected] Website: http://www.academicjournals.org/journal/AJHC Submit manuscript online http://ms.academicjournals.me/. Editorial Board Dr. Antonio J. Monroy Antón Dr Jephias Mapuva Department of Business Economics African Centre for Citizenship and Democracy Universidad Carlos III , [ACCEDE];School of Government; University of the Western Cape, Madrid, Spain. -

Temples and Tombs Treasures of Egyptian Art from the British Museum
Temples and Tombs Treasures of Egyptian Art from The British Museum Resource for Educators this is max size of image at 200 dpi; the sil is low res and for the comp only. if approved, needs to be redone carefully American Federation of Arts Temples and Tombs Treasures of Egyptian Art from The British Museum Resource for Educators American Federation of Arts © 2006 American Federation of Arts Temples and Tombs: Treasures of Egyptian Art from the British Museum is organized by the American Federation of Arts and The British Museum. All materials included in this resource may be reproduced for educational American Federation of Arts purposes. 212.988.7700 800.232.0270 The AFA is a nonprofit institution that organizes art exhibitions for presen- www.afaweb.org tation in museums around the world, publishes exhibition catalogues, and interim address: develops education programs. 122 East 42nd Street, Suite 1514 New York, NY 10168 after April 1, 2007: 305 East 47th Street New York, NY 10017 Please direct questions about this resource to: Suzanne Elder Burke Director of Education American Federation of Arts 212.988.7700 x26 [email protected] Exhibition Itinerary to Date Oklahoma City Museum of Art Oklahoma City, Oklahoma September 7–November 26, 2006 The Cummer Museum of Art and Gardens Jacksonville, Florida December 22, 2006–March 18, 2007 North Carolina Museum of Art Raleigh, North Carolina April 15–July 8, 2007 Albuquerque Museum of Art and History Albuquerque, New Mexico November 16, 2007–February 10, 2008 Fresno Metropolitan Museum of Art, History and Science Fresno, California March 7–June 1, 2008 Design/Production: Susan E. -

Victoria En Nubia
^J ^K FEBRERO/ MARZO 1980 7 francos (España: 150 pesetas) El Correo*la unesco 1a3 » Victoria Mí en IMubia fe 4.000 años de historia '/, salvados de las aguas ^W^W^Íi I t Foto Ù- Museo Nacional de Varsovia Caballo nubio TESOROS Este caballo pertenece a una pintura mural de la catedral de Faras que actualmente se con¬ DEL ARTE serva en el Museo de Varsovia como regalo del Gobierno sudanés. (Los gobiernos de Sudón y de Egipto donaron a cada uno de los países que participaron en la campaña de Nubia una parte MUNDIAL de los tesoros descubiertos por sus misiones arqueológicas). Faras, o Pachoras, según su nom¬ bre preérabe, era un importante centro de la Nubia sudanesa, situado muy cerca de la frontera con Egipto. De su excavación se encargó la expedición arqueológica polaca. El sitio fue uno de ^% los primeros de la Nubia sudanesa en quedar sumergidos por las aguas de la gran presa de Asuán. La catedral de Faras tuvo gran importancia en la historia del cristianismo en la Nubia inferior. En esta pintura el artista del siglo XII imita perfectamente los movimientos del caballo Sudán que se encabrita. Los dos apéndices bajo la pata delantera derecha parecen indicar que lo aquí reproducido era un ornamento que podía clavarse o colgarse. páginas ei Correo ^e ^a unesc° "LA HISTORIA DE LOS HOMBRES.. por Amadou-Mahtar M'Bow Una ventana abierta al mundo Lanzada por la Unesco, la mayor operación de salvamento arqueológico de todos los tiempos (1960-1980) FEBRERO-MARZO 1979 AÑO XXXII 5 VICTORIA EN NUBIA: EGIPTO por Shehata Adam Mohamed PUBLICADO EN 20 IDIOMAS 16 VICTORIA EN NUBIA: SUDAN por Negm-EI-Dln Mohamed Sherif Español Italiano Turco Inglés Hindi Urdu 14 MONUMENTOS SALVADOS DEL NILO Mapa Francés Tamul Catalán Ruso Hebreo Malayo 20 NUBIA REDESCUBIERTA Alemán Persa Coreano De la prehistoria a los tiempos faraónicos por Torgny Säve-Söderbergh Arabe Portugués Swahili Japonés Neerlandés 25 VICISITUDES DE UNA HISTORIA Del Imperio de Kush al Islam por William Y. -

Sudan a Country Study.Pdf
A Country Study: Sudan An Nilain Mosque, at the site of the confluence of the Blue Nile and White Nile in Khartoum Federal Research Division Library of Congress Edited by Helen Chapin Metz Research Completed June 1991 Table of Contents Foreword Acknowledgements Preface Country Profile Country Geography Society Economy Transportation Government and Politics National Security Introduction Chapter 1 - Historical Setting (Thomas Ofcansky) Early History Cush Meroe Christian Nubia The Coming of Islam The Arabs The Decline of Christian Nubia The Rule of the Kashif The Funj The Fur The Turkiyah, 1821-85 The Mahdiyah, 1884-98 The Khalifa Reconquest of Sudan The Anglo-Egyptian Condominium, 1899-1955 Britain's Southern Policy Rise of Sudanese Nationalism The Road to Independence The South and the Unity of Sudan Independent Sudan The Politics of Independence The Abbud Military Government, 1958-64 Return to Civilian Rule, 1964-69 The Nimeiri Era, 1969-85 Revolutionary Command Council The Southern Problem Political Developments National Reconciliation The Transitional Military Council Sadiq Al Mahdi and Coalition Governments Chapter 2 - The Society and its Environment (Robert O. Collins) Physical Setting Geographical Regions Soils Hydrology Climate Population Ethnicity Language Ethnic Groups The Muslim Peoples Non-Muslim Peoples Migration Regionalism and Ethnicity The Social Order Northern Arabized Communities Southern Communities Urban and National Elites Women and the Family Religious -

Two Firing Structures from Ancient Sudan: an Archaeological Note
Dotawo: A Journal of Nubian Studies Volume 3 Know-Hows and Techniques in Ancient Sudan Article 3 2016 Two Firing Structures from Ancient Sudan: An Archaeological Note Sebastien Maillot [email protected] Follow this and additional works at: https://digitalcommons.fairfield.edu/djns Recommended Citation Maillot, Sebastien (2016) "Two Firing Structures from Ancient Sudan: An Archaeological Note," Dotawo: A Journal of Nubian Studies: Vol. 3 , Article 3. Available at: https://digitalcommons.fairfield.edu/djns/vol3/iss1/3 This item has been accepted for inclusion in DigitalCommons@Fairfield by an authorized administrator of DigitalCommons@Fairfield. It is brought to you by DigitalCommons@Fairfield with permission from the rights- holder(s) and is protected by copyright and/or related rights. You are free to use this item in any way that is permitted by the copyright and related rights legislation that applies to your use. For other uses, you need to obtain permission from the rights-holder(s) directly, unless additional rights are indicated by a Creative Commons license in the record and/or on the work itself. For more information, please contact [email protected]. 41 Two Firing Structures from Ancient Sudan: An Archaeological Note Sébastien Maillot* The Kushite temples of Amun at Dukki Gel1 and Dangeil2 (fig. 1) fea- ture in their precincts large heaps of ash mixed with ceramic coni- cal moulds (or “bread moulds”), which are dumps associated with the production of food offerings for the cult. Excavation of the areas dedicated to these activities resumed in 2013 at both sites, uncover- ing numerous firing and storage features. -
A Brief Description of the Nobiin Language and History by Nubantood Khalil, (Nubian Language Society)
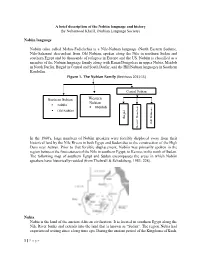
A brief description of the Nobiin language and history By Nubantood Khalil, (Nubian Language Society) Nobiin language Nobiin (also called Mahas-Fadichcha) is a Nile-Nubian language (North Eastern Sudanic, Nilo-Saharan) descendent from Old Nubian, spoken along the Nile in northern Sudan and southern Egypt and by thousands of refugees in Europe and the US. Nobiin is classified as a member of the Nubian language family along with Kenzi/Dongolese in upper Nubia, Meidob in North Darfur, Birgid in Central and South Darfur, and the Hill Nubian languages in Southern Kordofan. Figure 1. The Nubian Family (Bechhaus 2011:15) Central Nubian Western Northern Nubian Nubian ▪ Nobiin ▪ Meidob ▪ Old Nubian Birgid Hill Nubians Kenzi/ Donglese In the 1960's, large numbers of Nobiin speakers were forcibly displaced away from their historical land by the Nile Rivers in both Egypt and Sudan due to the construction of the High Dam near Aswan. Prior to that forcible displacement, Nobiin was primarily spoken in the region between the first cataract of the Nile in southern Egypt, to Kerma, in the north of Sudan. The following map of southern Egypt and Sudan encompasses the areas in which Nobiin speakers have historically resided (from Thelwall & Schadeberg, 1983: 228). Nubia Nubia is the land of the ancient African civilization. It is located in southern Egypt along the Nile River banks and extends into the land that is known as “Sudan”. The region Nubia had experienced writing since a long time ago. During the ancient period of the Kingdoms of Kush, 1 | P a g e the Kushite/Nubians used the hieroglyphic writing system. -

Amarna Period Down to the Opening of Sety I's Reign
oi.uchicago.edu STUDIES IN ANCIENT ORIENTAL CIVILIZATION * NO.42 THE ORIENTAL INSTITUTE OF THE UNIVERSITY OF CHICAGO Thomas A. Holland * Editor with the assistance of Thomas G. Urban oi.uchicago.edu oi.uchicago.edu Internet publication of this work was made possible with the generous support of Misty and Lewis Gruber THE ROAD TO KADESH A HISTORICAL INTERPRETATION OF THE BATTLE RELIEFS OF KING SETY I AT KARNAK SECOND EDITION REVISED WILLIAM J. MURNANE THE ORIENTAL INSTITUTE OF THE UNIVERSITY OF CHICAGO STUDIES IN ANCIENT ORIENTAL CIVILIZATION . NO.42 CHICAGO * ILLINOIS oi.uchicago.edu Library of Congress Catalog Card Number: 90-63725 ISBN: 0-918986-67-2 ISSN: 0081-7554 The Oriental Institute, Chicago © 1985, 1990 by The University of Chicago. All rights reserved. Published 1990. Printed in the United States of America. oi.uchicago.edu TABLE OF CONTENTS List of M aps ................................ ................................. ................................. vi Preface to the Second Edition ................................................................................................. vii Preface to the First Edition ................................................................................................. ix List of Bibliographic Abbreviations ..................................... ....................... xi Chapter 1. Egypt's Relations with Hatti From the Amarna Period Down to the Opening of Sety I's Reign ...................................................................... ......................... 1 The Clash of Empires -

Graffiti-As-Devotion.Pdf
lsa.umich.edu/kelsey/ i lsa.umich.edu/kelsey/ lsa.umich.edu/kelsey/ iii Edited by Geoff Emberling and Suzanne Davis Along the Nile and Beyond Kelsey Museum Publication 16 Kelsey Museum of Archaeology University of Michigan, 2019 lsa.umich.edu/kelsey/ iv Graffiti as Devotion along the Nile and Beyond The Kelsey Museum of Archaeology, Ann Arbor 48109 © 2019 by The Kelsey Museum of Archaeology and the individual authors All rights reserved Published 2019 ISBN-13: 978-0-9906623-9-6 Library of Congress Control Number: 2019944110 Kelsey Museum Publication 16 Series Editor Leslie Schramer Cover design by Eric Campbell This book was published in conjunction with the special exhibition Graffiti as Devotion along the Nile: El-Kurru, Sudan, held at the Kelsey Museum of Archaeology in Ann Arbor, Michigan. The exhibition, curated by Geoff Emberling and Suzanne Davis, was on view from 23 August 2019 through 29 March 2020. An online version of the exhibition can be viewed at http://exhibitions.kelsey.lsa.umich.edu/graffiti-el-kurru Funding for this publication was provided by the University of Michigan College of Literature, Science, and the Arts and the University of Michigan Office of Research. This book is available direct from ISD Book Distributors: 70 Enterprise Drive, Suite 2 Bristol, CT 06010, USA Telephone: (860) 584-6546 Email: [email protected] Web: www.isdistribution.com A PDF is available for free download at https://lsa.umich.edu/kelsey/publications.html Printed in South Korea by Four Colour Print Group, Louisville, Kentucky. ♾ This paper meets the requirements of ANSI/NISO Z39.48-1992 (Permanence of Paper). -

Baqt Treaty | 1 BAQT TREATY
Baqt Treaty | 1 BAQT TREATY The Baqt was a negotiated agreement between ‘Abdallah ibn Sa’id ibn Abi Sarh, the Umayyad governor of Egypt, and the Nubian king of MAKOURIA. It was concluded at DONGOLA in A.D. 632 following an unsuccessful Muslim attempt to subjugate the Nubian kingdom. The name Baqt is presumed to be derivative from Greek pakton (agreement), and is unique to this one document; no other covenant made by the medieval Arabs was given the same designation. This reflects the fact that the agreement itself was in many ways unique in the annals of Arab foreign relations, since it exempted the Nubians from the dar al-harb (the community of nations at war with Islam) without including them in the dar al-Islam (the community of Islam). Several different versions of the Baqt have been recorded by different authors. Most of them wrote at a time when the agreement was still nominally in force but long after the original date of its execution. The earliest written account of the Baqt appears to be that of Al-Baladhuri (d. 892), who merely states that the Nubians were exempted from paying JIZYAH (poll tax) but agreed to furnish annually a gift of 300 slaves in exchange for the equivalent value in food. Later writers mention various other conditions, such as provision of horses by the Nubians and of wine and various kinds of cloth by the Egyptians. The fullest recorded version of the agreement is that in al-MAQRIZI’s al-Khitat (Plans), written more than 700 years after the actual date of the treaty. -

The PCMA UW Research Centre in Cairo: 60 Years in the Field
The PCMA UW Research Centre in Cairo: 60 years in the field “To be a player in world archaeology, we must be in Egypt!” Prof. Kazimierz Michałowski The indisputed position of Polish Mediterranean archae- ology and Polish archaeologists in Egypt is the effect of dozens of years of excavations at different archaeological sites as well as outstanding research, countless publica- tions and many impressive and inspiring projects. The beginnings, however, were quite modest. More than 80 years ago Professor Kazimierz Michałowski, representing the University of Warsaw, joined forces with the French Institute of Oriental Archaeology (IFAO) in Cairo to excavate at Edfu. Pooling resources and organizational skills, the two institutions carried out three season of fieldwork, opening the field of international stud- ies of ancient cultures to Polish archaeologists. These three seasons—before the outbreak of World War II put a stop to further investigations—were of immeasurable significance for Polish culture and science. Since that memorable year, 1937, Polish archaeological presence in the ancient Mediter- ranean is an unquestioned fact. An archaeological return to the world of ancient civilizations was Michałowski’s main objective after the war. First were sites on the Black Sea (Myrmekion, Olbia, Kalos Limen) in the mid-1950s, then Novae, Renata Kucharczyk a Roman fortress on the Danube. Polish archaeologists Polish Centre of Mediterranean soon launched research in Egypt, appearing from this Archaeology, University of Warsaw Polish Archaeology in the Mediterranean 28/2 PAM 28/2 (2019) Kucharczyk 2019: 15–20 DOI: 10.31338/uw.2083-537X.pam28.2.01 EGYPT The PCMA UW Research Centre in the field: 60 years of fieldwork and research point on as an important independent a major international project launched by partner of the Egyptian Ministry of An- UNESCO and headed by Michałowski. -

120 'NOTITIAE Fouilles Et Travaux Au Soudan, 1955-1960 (*) J. LECLANT
120 'NOTITIAE Fouilles et travaux au Soudan, 1955-1960 (*) J. LECLANT - Strasbourg En juin 1955, M. Jean Vercoutter a remplacé M. P. L. Shinnie comme « Comm.issioner for Archaeology » à la tête du Service des Anti quités du Soudan('). La tâche essentielle qui s'est posée à lui a été l'étude et la sauvegarde des sites et monuments situés dans la zone menacée de submersion par la construction du Sadd el Ah; comme on le sait,, la vallée, au Soudan, (*) Pour les fouilles et travaux au Soudan depuis 1948, cf. Orientalia, 20, 1951, p. 351-355; 22, 1953, p. 105; 24, 1955, p. 159-163, ainsi que J. Le clatit, French Archaeological Digest, Archaeology 1945-1955, Part I (New York, 1956), p. 73-74. (I) Les importants travaux de J. Vercoutter et de ses collaborateurs Thabit Hassan Thabit, Sadik Nur, Nigm ed Din Mohammed, Aluned Hassan Ibrahim ont été l'objet de rapports très réguliers et précis, pré sentés par M. J. Vercoutter. Ils sont à la base du présent compte rendu, essntieh1ement bibliographique. - 10 Il y a d'abord les rapports du Ser vice des Antiquités du Soudan, que nous citerons sous la forme abrégée Report, suivi de la date de la campagne envisagée; afin de permettre un report bibliographique exact au titre complet de ces plaquettes, nous indiquons ici-même leurs titres: Sudan Government, Report on the Antiq«i ties Service and Museums 1954-1955, by H. N. Chittick (McCorquodale and Co., Sudan, s. d.); The Republic of the Sudan, Report on the Antiquities Service and Museums 1955-1956, by Dr.