Rates of Molecular Evolution and Diversification in Plants: Chloroplast
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Method to Estimate Dry-Kiln Schedules and Species Groupings: Tropical and Temperate Hardwoods
United States Department of Agriculture Method to Estimate Forest Service Forest Dry-Kiln Schedules Products Laboratory Research and Species Groupings Paper FPL–RP–548 Tropical and Temperate Hardwoods William T. Simpson Abstract Contents Dry-kiln schedules have been developed for many wood Page species. However, one problem is that many, especially tropical species, have no recommended schedule. Another Introduction................................................................1 problem in drying tropical species is the lack of a way to Estimation of Kiln Schedules.........................................1 group them when it is impractical to fill a kiln with a single Background .............................................................1 species. This report investigates the possibility of estimating kiln schedules and grouping species for drying using basic Related Research...................................................1 specific gravity as the primary variable for prediction and grouping. In this study, kiln schedules were estimated by Current Kiln Schedules ..........................................1 establishing least squares relationships between schedule Method of Schedule Estimation...................................2 parameters and basic specific gravity. These relationships were then applied to estimate schedules for 3,237 species Estimation of Initial Conditions ..............................2 from Africa, Asia and Oceana, and Latin America. Nine drying groups were established, based on intervals of specific Estimation -

NSW Rainforest Trees Part
This document has been scanned from hard-copy archives for research and study purposes. Please note not all information may be current. We have tried, in preparing this copy, to make the content accessible to the widest possible audience but in some cases we recognise that the automatic text recognition maybe inadequate and we apologise in advance for any inconvenience this may cause. · RESEARCH NOTE No. 35 ~.I~=1 FORESTRY COMMISSION OF N.S.W. RESEARCH NOTE No. 35 P)JBLISHED 197R N.S.W. RAINFOREST TREES PART VII FAMILIES: PROTEACEAE SANTALACEAE NYCTAGINACEAE GYROSTEMONACEAE ANNONACEAE EUPOMATIACEAE MONIMIACEAE AUTHOR A.G.FLOYD (Research Note No. 35) National Library of Australia card number and ISBN ISBN 0 7240 13997 ISSN 0085-3984 INTRODUCTION This is the seventh in a series ofresearch notes describing the rainforest trees of N.S. W. Previous publications are:- Research Note No. 3 (I 960)-N.S.W. Rainforest Trees. Part I Family LAURACEAE. A. G. Floyd and H. C. Hayes. Research Note No. 7 (1961)-N.S.W. Rainforest Trees. Part II Families Capparidaceae, Escalloniaceae, Pittosporaceae, Cunoniaceae, Davidsoniaceae. A. G. Floyd and H. C. Hayes. Research Note No. 28 (I 973)-N.S.W. Rainforest Trees. Part III Family Myrtaceae. A. G. Floyd. Research Note No. 29 (I 976)-N.S.W. Rainforest Trees. Part IV Family Rutaceae. A. G. Floyd. Research Note No. 32 (I977)-N.S.W. Rainforest Trees. Part V Families Sapindaceae, Akaniaceae. A. G. Floyd. Research Note No. 34 (1977)-N.S.W. Rainforest Trees. Part VI Families Podocarpaceae, Araucariaceae, Cupressaceae, Fagaceae, Ulmaceae, Moraceae, Urticaceae. -

Tasmanian Wilderness World Heritage Area
Appendix 4 1 World Heritage Values of the Tasmanian Wilderness 1.1 Note that the Department of the Environment's website states that: A draft Statement of Outstanding Universal Value which will take into account the new areas added in 2013 is expected to be considered by the World Heritage Committee in 2014. Outstanding Universal Value 1.2 The Tasmanian Wilderness is an extensive, wild, beautiful temperate land where cultural heritage of the Tasmanian Aboriginal people is preserved. 1.3 It is one of the three largest temperate wilderness areas remaining in the Southern Hemisphere. The region is home to some of the deepest and longest caves in Australia. It is renowned for its diversity of flora, and some of the longest lived trees and tallest flowering plants in the world grow in the area. The Tasmanian Wilderness is a stronghold for several animals that are either extinct or threatened on mainland Australia. 1.4 In the southwest Aboriginal people developed a unique cultural tradition based on a specialized stone and bone toolkit that enabled the hunting and processing of a single prey species (Bennett's wallaby) that provided nearly all of their dietary protein and fat. Extensive limestone cave systems contain rock art sites that have been dated to the end of the Pleistocene period. Southwest Tasmanian Aboriginal artistic expression during the last Ice Age is only known from the dark recesses of limestone caves. 1.5 The Tasmanian Wilderness was inscribed on the World Heritage List in 1982 and extended in 1989, 2010, 2012 and again in 2013. -

The Macadamia Industry in New Zealand
Copyright is owned by the Author of the thesis. Pennission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere without the pennission of the Author. THE MACADAMIA INDUSTRY IN NEW ZEALAND A Thesis for the Degree of Master of Philosophy At Massey University Palmerston North Bernard Coleman Whangarei 2005 ABSTRACT The New Zealand macadamia industry has been characterised by many small plantings, lifestyle blocks up to 1500 trees and two commercial plantations with more than 10000 trees. Completed research programmes have been few, mainly because government funding in horticulture has been channelled to the needs of the major crops such as kiwifruit. Changes in political policy affected funding for minor horticultural crops and spasmodic cuts in finance severely hindered long-term research projects. Because of its small size the macadamia industry had limited funds available from members but some research programmes have been completed including pest control, tree nutrition, basal stain and future research needs. A private consultant, Ian Gordon has carried out variety trials on a local selection. Several selections have been planted in different locations and have proven to be useful in pollination of Beaumont, the main variety planted in New Zealand. Within the limits, set by climatic factors, the suitable growing areas are north of a line from New Plymouth to Gisbome. Both of those areas are marginal for commercial planting but sites on the sheltered north facing positions could grow satisfactory yields. Yields per tree, generally have been below commercial requirements. -
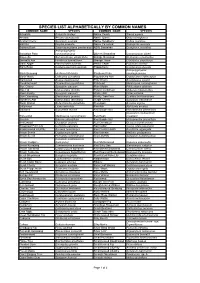
Species List Alphabetically by Common Names
SPECIES LIST ALPHABETICALLY BY COMMON NAMES COMMON NAME SPECIES COMMON NAME SPECIES Actephila Actephila lindleyi Native Peach Trema aspera Ancana Ancana stenopetala Native Quince Guioa semiglauca Austral Cherry Syzygium australe Native Raspberry Rubus rosifolius Ball Nut Floydia praealta Native Tamarind Diploglottis australis Banana Bush Tabernaemontana pandacaqui NSW Sassafras Doryphora sassafras Archontophoenix Bangalow Palm cunninghamiana Oliver's Sassafras Cinnamomum oliveri Bauerella Sarcomelicope simplicifolia Orange Boxwood Denhamia celastroides Bennetts Ash Flindersia bennettiana Orange Thorn Citriobatus pauciflorus Black Apple Planchonella australis Pencil Cedar Polyscias murrayi Black Bean Castanospermum australe Pepperberry Cryptocarya obovata Archontophoenix Black Booyong Heritiera trifoliolata Picabeen Palm cunninghamiana Black Wattle Callicoma serratifolia Pigeonberry Ash Cryptocarya erythroxylon Blackwood Acacia melanoxylon Pink Cherry Austrobuxus swainii Bleeding Heart Omalanthus populifolius Pinkheart Medicosma cunninghamii Blue Cherry Syzygium oleosum Plum Myrtle Pilidiostigma glabrum Blue Fig Elaeocarpus grandis Poison Corkwood Duboisia myoporoides Blue Lillypilly Syzygium oleosum Prickly Ash Orites excelsa Blue Quandong Elaeocarpus grandis Prickly Tree Fern Cyathea leichhardtiana Blueberry Ash Elaeocarpus reticulatus Purple Cherry Syzygium crebrinerve Blush Walnut Beilschmiedia obtusifolia Red Apple Acmena ingens Bollywood Litsea reticulata Red Ash Alphitonia excelsa Bolwarra Eupomatia laurina Red Bauple Nut Hicksbeachia -

The Macadamia Industry in New Zealand
Copyright is owned by the Author of the thesis. Pennission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere without the pennission of the Author. THE MACADAMIA INDUSTRY IN NEW ZEALAND A Thesis for the Degree of Master of Philosophy At Massey University Palmerston North Bernard Coleman Whangarei 2005 ABSTRACT The New Zealand macadamia industry has been characterised by many small plantings, lifestyle blocks up to 1500 trees and two commercial plantations with more than 10000 trees. Completed research programmes have been few, mainly because government funding in horticulture has been channelled to the needs of the major crops such as kiwifruit. Changes in political policy affected funding for minor horticultural crops and spasmodic cuts in finance severely hindered long-term research projects. Because of its small size the macadamia industry had limited funds available from members but some research programmes have been completed including pest control, tree nutrition, basal stain and future research needs. A private consultant, Ian Gordon has carried out variety trials on a local selection. Several selections have been planted in different locations and have proven to be useful in pollination of Beaumont, the main variety planted in New Zealand. Within the limits, set by climatic factors, the suitable growing areas are north of a line from New Plymouth to Gisbome. Both of those areas are marginal for commercial planting but sites on the sheltered north facing positions could grow satisfactory yields. Yields per tree, generally have been below commercial requirements. -

Rates of Molecular Evolution and Diversification in Plants: Chloroplast
Duchene and Bromham BMC Evolutionary Biology 2013, 13:65 http://www.biomedcentral.com/1471-2148/13/65 RESEARCH ARTICLE Open Access Rates of molecular evolution and diversification in plants: chloroplast substitution rates correlate with species-richness in the Proteaceae David Duchene* and Lindell Bromham Abstract Background: Many factors have been identified as correlates of the rate of molecular evolution, such as body size and generation length. Analysis of many molecular phylogenies has also revealed correlations between substitution rates and clade size, suggesting a link between rates of molecular evolution and the process of diversification. However, it is not known whether this relationship applies to all lineages and all sequences. Here, in order to investigate how widespread this phenomenon is, we investigate patterns of substitution in chloroplast genomes of the diverse angiosperm family Proteaceae. We used DNA sequences from six chloroplast genes (6278bp alignment with 62 taxa) to test for a correlation between diversification and the rate of substitutions. Results: Using phylogenetically-independent sister pairs, we show that species-rich lineages of Proteaceae tend to have significantly higher chloroplast substitution rates, for both synonymous and non-synonymous substitutions. Conclusions: We show that the rate of molecular evolution in chloroplast genomes is correlated with net diversification rates in this large plant family. We discuss the possible causes of this relationship, including molecular evolution driving diversification, speciation increasing the rate of substitutions, or a third factor causing an indirect link between molecular and diversification rates. The link between the synonymous substitution rate and clade size is consistent with a role for the mutation rate of chloroplasts driving the speed of reproductive isolation. -

Evolutionary History of Floral Key Innovations in Angiosperms Elisabeth Reyes
Evolutionary history of floral key innovations in angiosperms Elisabeth Reyes To cite this version: Elisabeth Reyes. Evolutionary history of floral key innovations in angiosperms. Botanics. Université Paris Saclay (COmUE), 2016. English. NNT : 2016SACLS489. tel-01443353 HAL Id: tel-01443353 https://tel.archives-ouvertes.fr/tel-01443353 Submitted on 23 Jan 2017 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. NNT : 2016SACLS489 THESE DE DOCTORAT DE L’UNIVERSITE PARIS-SACLAY, préparée à l’Université Paris-Sud ÉCOLE DOCTORALE N° 567 Sciences du Végétal : du Gène à l’Ecosystème Spécialité de Doctorat : Biologie Par Mme Elisabeth Reyes Evolutionary history of floral key innovations in angiosperms Thèse présentée et soutenue à Orsay, le 13 décembre 2016 : Composition du Jury : M. Ronse de Craene, Louis Directeur de recherche aux Jardins Rapporteur Botaniques Royaux d’Édimbourg M. Forest, Félix Directeur de recherche aux Jardins Rapporteur Botaniques Royaux de Kew Mme. Damerval, Catherine Directrice de recherche au Moulon Président du jury M. Lowry, Porter Curateur en chef aux Jardins Examinateur Botaniques du Missouri M. Haevermans, Thomas Maître de conférences au MNHN Examinateur Mme. Nadot, Sophie Professeur à l’Université Paris-Sud Directeur de thèse M. -

(OUV) of the Wet Tropics of Queensland World Heritage Area
Handout 2 Natural Heritage Criteria and the Attributes of Outstanding Universal Value (OUV) of the Wet Tropics of Queensland World Heritage Area The notes that follow were derived by deconstructing the original 1988 nomination document to identify the specific themes and attributes which have been recognised as contributing to the Outstanding Universal Value of the Wet Tropics. The notes also provide brief statements of justification for the specific examples provided in the nomination documentation. Steve Goosem, December 2012 Natural Heritage Criteria: (1) Outstanding examples representing the major stages in the earth’s evolutionary history Values: refers to the surviving taxa that are representative of eight ‘stages’ in the evolutionary history of the earth. Relict species and lineages are the elements of this World Heritage value. Attribute of OUV (a) The Age of the Pteridophytes Significance One of the most significant evolutionary events on this planet was the adaptation in the Palaeozoic Era of plants to life on the land. The earliest known (plant) forms were from the Silurian Period more than 400 million years ago. These were spore-producing plants which reached their greatest development 100 million years later during the Carboniferous Period. This stage of the earth’s evolutionary history, involving the proliferation of club mosses (lycopods) and ferns is commonly described as the Age of the Pteridophytes. The range of primitive relict genera representative of the major and most ancient evolutionary groups of pteridophytes occurring in the Wet Tropics is equalled only in the more extensive New Guinea rainforests that were once continuous with those of the listed area. -

Seed Dynamics and Seedling Establishment of Woody Species in the Tropical Savannas of Central Brazil (Cerrado) Ana A
University of Miami Scholarly Repository Open Access Dissertations Electronic Theses and Dissertations 2010-04-08 Seed Dynamics and Seedling Establishment of Woody Species in the Tropical Savannas of Central Brazil (Cerrado) Ana A. Salazar Parra University of Miami, [email protected] Follow this and additional works at: https://scholarlyrepository.miami.edu/oa_dissertations Recommended Citation Salazar Parra, Ana A., "Seed Dynamics and Seedling Establishment of Woody Species in the Tropical Savannas of Central Brazil (Cerrado)" (2010). Open Access Dissertations. 371. https://scholarlyrepository.miami.edu/oa_dissertations/371 This Open access is brought to you for free and open access by the Electronic Theses and Dissertations at Scholarly Repository. It has been accepted for inclusion in Open Access Dissertations by an authorized administrator of Scholarly Repository. For more information, please contact [email protected]. UNIVERSITY OF MIAMI SEED DYNAMICS AND SEEDLING ESTABLISHMENT OF WOODY SPECIES IN THE TROPICAL SAVANNAS OF CENTRAL BRAZIL (CERRADO) By Ana A. Salazar Parra A DISSERTATION Submitted to the Faculty of the University of Miami in partial fulfillment of the requirements for the degree of Doctor of Philosophy Coral Gables, Florida May 2010 ©2010 Ana A. Salazar Parra All Rights Reserved UNIVERSITY OF MIAMI A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy SEED DYNAMICS AND SEEDLING ESTABLISHMENT OF WOODY SPECIES IN THE TROPICAL SAVANNAS OF CENTRAL BRAZIL (CERRADO) Ana A. Salazar Parra Approved: ________________ _________________ Guillermo Goldstein, Ph.D. Terri A. Scandura, Ph.D. Professor of Biology Dean of the Graduate School ________________ _________________ David Janos, Ph.D. Leonel Sternberg, Ph.D. -

Toronia Toru
Toronia toru COMMON NAME Toru, toro, toto, mihimihi SYNONYMS Persoonia toru A.Cunn. FAMILY Proteaceae AUTHORITY Toronia toru (A.Cunn.) L.A.S.Johnson et B.G.Briggs FLORA CATEGORY Vascular – Native ENDEMIC TAXON Yes ENDEMIC GENUS Yes ENDEMIC FAMILY No STRUCTURAL CLASS Trees & Shrubs - Dicotyledons NVS CODE TORTOR Toronia toru. Photographer: Wayne Bennett CHROMOSOME NUMBER 2n = 28 CURRENT CONSERVATION STATUS 2012 | Not Threatened PREVIOUS CONSERVATION STATUSES 2009 | Not Threatened 2004 | Not Threatened BRIEF DESCRIPTION Small bushy tree with long narrow leathery smooth edged-leaves that are yellowish when young and red when dying. Leaves 16-20cm long by 8-15mm wide, sharp tipped. Flowers yellowish or white, small, in clusters. Fruit dark purple, 12-18mm long. DISTRIBUTION Endemic. North Island from Te Paki south to the mouth of the Waihaha River, on the western side of Lake Taupo. However south of Auckland, toru is mostly found in the east from the Coromandel throughout the Bay Kennedy Bay, November. Photographer: John of Plenty to about Atiamuri. In the western Waikato it is known locally Smith-Dodsworth from the northern end of the Aotea Harbour, near Kawhia (Rakaunui), Te Kauri and Whenuapo in the Taumatatotara Range HABITAT Coastal to montane mostly on infertile soils, in open shrubland (especially gumland), early successional forest and along ridge lines and around slip scars in kauri (Agathis australis) and/or tanekaha (Phyllocladus trichomanoides) dominated forest. It is locally abundant on silicic igneous rocks such as rhyolite, ignimbrite and pumice. FEATURES Small dioecious (or gynodioecious) tree up to 12 m tall; trunk 1 or more arising from base, 0.2-0.3 m dbh; bark firm (not flaking), grey, grey-brown, brown or mottled grey, grey-brown. -

Kermadecia Brinoniae (Proteaceae: Macadamieae), a New Species from New Caledonia Previously Confused with K
Kermadecia brinoniae (Proteaceae: Macadamieae), a new species from New Caledonia previously confused with K. elliptica Helen C.F. Hopkins & Yohan Pillon Abstract HOPKINS, H.C.F. & Y. PILLON (2019). Kermadecia brinoniae (Proteaceae: Macadamieae), a new species from New Caledonia previously confused with K. elliptica. In English, English and French abstracts. Candollea 74: 85 – 92. DOI: http://dx.doi.org/10.15553/c2019v741a9 Kermadecia brinoniae H.C. Hopkins & Pillon (Proteaceae), which occurs in southern New Caledonia, principally on ultramafic substrates, is described and illustrated. Material of this species was previously included in Kermadecia elliptica Brongn. & Gris. An amended description is given for the latter and a lectotype designated. These two species differ in the prominence of the venation on the lower leaf surface and in their ecology. Kermadecia brinoniae, like other species of Proteaceae growing on ultramafic substrates of New Caledonia, has a relatively high content of manganese in its leaves. A revised key to the species of Kermadecia Brongn. & Gris is provided as well as notes on the morphology of the pollen presenter. Résumé HOPKINS, H. C. F. & Y. PILLON (2019). Kermadecia brinoniae (Proteaceae: Macadamieae), une nouvelle espèce de Nouvelle-Calédonie auparavant confondue avec K. elliptica. En anglais, résumés anglais et français. Candollea 74: 85 – 92. DOI: http://dx.doi.org/10.15553/c2019v741a9 Kermadecia brinoniae H.C. Hopkins & Pillon (Proteaceae), du sud de la Nouvelle-Calédonie, essentiellement sur subs- trat ultramafique, est ici décrite et illustrée. Les spécimens de cette espèce ayant été auparavant inclus dans Kermadecia elliptica Brongn. & Gris, la description de ce taxon est revue et un lectotype est désigné.