Thermal Relations 10
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Development and Structure of the Excretory System the Nephron
Biology 224 Human Anatomy and Physiology II Week 7; Lecture 1; Monday Dr. Stuart S. Sumida Development and Structure of the Excretory System The Nephron DEVELOPMENT AND STRUCTURE OF EXCRETORY SYSTEM EXCRETORY SYSTEM REVIEW • Kidneys derived from INTERMEDIATE MESODERM. • Kidney starts out as a SEGMENTAL STRUCTURE. • Bladder, as part of embryonic gut tube: lining derived from endoderm. • Note, this is EXCRETION, NOT “ELIMINATION.” EARLY KIDNEY DEVELOPMENT • There is a segment of intermediate mesoderm for every body segment. • Earliest kidney appears in the cervical region of the body! (About week 3.) Called the PRONEPHROS. • Develop very close to the gonads. (They battle it out for the nearby ducts.) THE PRONEPHROS • The early anteriorly developed kidney parts. • Has NO EXCRETORY FUNCTION. • Functions to INDUCE DEVELOPMENT of middle segments of intermediate meosderm into the MESONEPHROS. THE MESONEPHROS • Some think it is the functioning embryonic kidney. Some think is has no excretory function. • WE DO KNOW that the duct that attaches to it (THE MESONEPHRIC DUCT) is very important in INDUCING DEVELOPMENT OF THE CAUDAL KIDNEY SEGMENTS (the METANEPHROS). Mesonephric Duct reaches all the way to end of gut tube (cloaca). We need to... • Attach the ducts to the hindend kidney (the metanephros). • Split the bladder away from the gut tube. After the mesonephric duct attaches to cloaca, the embryonic URETER grows from caudal to cranial to attach to mass of metanephric kidney. A septum, the URORECTAL SEPTUM, grows between the more dorsal part of the gut tube and the more ventral part that will become the bladder. Note that attached to the bladder are right and left ueters, the allantois (an extraembryonic membrane). -

The Contribution of Nasal Countercurrent Heat Exchange to Water Balance in the Northern Elephant Seal, Mirounga Angustirostris
J. exp. Biol. 113, 447-454 (1984) 447 Printed in Great Britain © The Company of Biologists Limited 1984 THE CONTRIBUTION OF NASAL COUNTERCURRENT HEAT EXCHANGE TO WATER BALANCE IN THE NORTHERN ELEPHANT SEAL, MIROUNGA ANGUSTIROSTRIS BY ANTHONY C. HUNTLEY, DANIEL P. COSTA Long Marine Laboratory, Center for Marine Studies, University of California, Santa Cruz, U.SA. AND ROBERT D. RUBIN Department of Life Sciences, Santa Rosa Junior College, Santa Rosa, California, U.SA Accepted 29 May 1984 SUMMARY 1. Elephant seals fast completely from food and water for 1-3 months during terrestrial breeding. Temporal countercurrent heat exchange in the nasal passage reduces expired air temperature (Te) below body temperature (Tb)- 2. At a mean ambient temperature of 13'7°C, Te is 20*9 °C. This results in the recovery of 71-5 % of the water added to inspired air. 3. The amount of cooling of the expired air (Tb~Te) and the percentage of water recovery varies inversely with ambient temperature. 4. Total nasal surface area available for heat and water exchange, located in the highly convoluted nasal turbinates, is estimated to be 720 cm2 in weaned pups and 3140 cm2 in an adult male. 5. Nasal temporal countercurrent heat exchange reduces total water loss sufficiently to allow maintenance of water balance using metabolic water production alone. INTRODUCTION Northern elephant seals, Mirounga angustirostis (Gill), are exceptional among pin- nipeds in the duration of their terrestrial breeding fast. During this time, they volun- tarily forgo both food and water while remaining active on the rookery. The length of these fasts varies with age, sex and social status. -

(Somniosus Pacificus) Predation on Steller Sea Lions (Eumetopias
297 Abstract—Temperature data re- In cold blood: evidence of Pacific sleeper shark ceived post mortem in 2008–13 from 15 of 36 juvenile Steller sea (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus) that lions (Eumetopias jubatus) in the Gulf of Alaska had been surgically implanted in 2005–11 with dual life history 1,2 transmitters (LHX tags) indicat- Markus Horning (contact author) ed that all 15 animals died by Jo-Ann E. Mellish3,4 predation. In 3 of those 15 cases, at least 1 of the 2 LHX tags was Email address for contact author: [email protected] ingested by a cold-blooded pred- ator, and those tags recorded, 1 Department of Fisheries and Wildlife and 3 Alaska Sea Life Center immediately after the sea lion’s Marine Mammal Institute P.O. Box 1329 death, temperatures that cor- College of Agricultural Sciences Seward, Alaska 99664 responded to deepwater values. Oregon State University 4 School of Fisheries and Ocean Sciences 2030 SE Marine Science Drive These tags were regurgitated or University of Alaska Fairbanks Newport, Oregon 97365 passed 5–11 days later by preda- P.O. Box 757220 2 tors. Once they sensed light and Coastal Oregon Marine Experiment Station Fairbanks, Alaska 99755-7220 Oregon State University air, the tags commenced trans- 2030 SE Marine Science Drive missions as they floated at the Newport, Oregon 97365 ocean surface, reporting tem- peratures that corresponded to regional sea-surface estimates. The circumstances related to the tag in a fourth case were ambig- The western distinct population seg- mercial fisheries (National Research uous. -

Determinants and Consequences of Interspeciwc Body Size Variation in Tetraphyllidean Tapeworms
Oecologia (2009) 161:759–769 DOI 10.1007/s00442-009-1410-1 COMMUNITY ECOLOGY - ORIGINAL PAPER Determinants and consequences of interspeciWc body size variation in tetraphyllidean tapeworms Haseeb Sajjad Randhawa · Robert Poulin Received: 3 December 2008 / Accepted: 20 June 2009 / Published online: 10 July 2009 © Springer-Verlag 2009 Abstract Tetraphyllidean cestodes are cosmopolitan, are relatively important variables in models explaining remarkably host speciWc, and form the most speciose and interspeciWc variations in tetraphyllidean tapeworm length. diverse group of helminths infecting elasmobranchs In addition, a negative relationship between tetraphyllidean (sharks, skates and rays). They show substantial interspe- body size and intensity of infection was apparent. These ciWc variation in a variety of morphological traits, including results suggest that space constraints and ambient tempera- body size. Tetraphyllideans represent therefore, an ideal ture, via their eVects on metabolism and growth, determine group in which to examine the relationship between para- adult tetraphyllidean cestode size. Consequently, a trade-oV site body size and abundance. The individual and combined between size and numbers is possibly imposed by external eVects of host size, environmental temperature, host habi- forces inXuencing host size, hence limiting physical space tat, host environment, host physiology, and host type (all or other resources available to the parasites. likely correlates of parasite body size) on parasite length were assessed using general linear model analyses using Keywords Tetraphyllidea · Abundance · Body size · data from 515 tetraphyllidean cestode species (182 species Depth · Latitude were included in analyses). The relationships between tetra- phyllidean cestode length and intensity and abundance of infection were assessed using simple linear regression anal- Introduction yses. -

Modeling and Exploiting Microbial Temperature Response
processes Review Modeling and Exploiting Microbial Temperature Response Philipp Noll, Lars Lilge, Rudolf Hausmann and Marius Henkel * Institute of Food Science and Biotechnology, Department of Bioprocess Engineering (150k), University of Hohenheim, Fruwirthstr. 12, 70599 Stuttgart, Germany; [email protected] (P.N.); [email protected] (L.L.); [email protected] (R.H.) * Correspondence: [email protected] Received: 15 December 2019; Accepted: 14 January 2020; Published: 17 January 2020 Abstract: Temperature is an important parameter in bioprocesses, influencing the structure and functionality of almost every biomolecule, as well as affecting metabolic reaction rates. In industrial biotechnology, the temperature is usually tightly controlled at an optimum value. Smart variation of the temperature to optimize the performance of a bioprocess brings about multiple complex and interconnected metabolic changes and is so far only rarely applied. Mathematical descriptions and models facilitate a reduction in complexity, as well as an understanding, of these interconnections. Starting in the 19th century with the “primal” temperature model of Svante Arrhenius, a variety of models have evolved over time to describe growth and enzymatic reaction rates as functions of temperature. Data-driven empirical approaches, as well as complex mechanistic models based on thermodynamic knowledge of biomolecular behavior at different temperatures, have been developed. Even though underlying biological mechanisms and mathematical models have been well-described, temperature as a control variable is only scarcely applied in bioprocess engineering, and as a conclusion, an exploitation strategy merging both in context has not yet been established. In this review, the most important models for physiological, biochemical, and physical properties governed by temperature are presented and discussed, along with application perspectives. -

Renal Physiology Background
RENAL PHYSIOLOGY Florida State University Advanced Topics in Biomedical Mathematics MAP5932, Spring 2007 03/16/07 Brinda Pamulapati Goal 1 Background of the kidney 2 Glomerulus 3 Mathematical model of Glomerulus 4 Co-current and counter- current mechanism 5 Mathematical model of the co-current and counter-current mechanism Kidney Kidney and Nephron picture Glomerulus and Bowman's Capsule Mathematical model of the glomerular filter There are 3 pressures that effect the rate of glomerular filtration: 1 the pressure inside the glomerular capillaries that promote filteration(p1) 2 the pressure inside the Bowman's capsule that opposes filtration(p2) 3 the colloidal osmotic pressure of the plasma proteins inside the capillaries that opposes filteration(pi) Schematic diagram of the glomerular filtration(one dimentional) q2 Qi q1 Qe x=0 x=L Mathematical model of the glomerular filter(cont.) dq 1 = K (P " P + ! ) dx f 2 1 c P1 ,P2 = hydrostatic pressure ! c = osmotic pressure of the suspended protei ns and formed elements of the blood K f = capillary filteration rate where osmotic pressure is ! c = RTc Conservation Equation ciQi = cq1 ! c ciQi = q1 ( from osmosis RTc) RT ! c = ! c = RTciQi / q1 Qi ! c = ! i (where ! i = RTci ) q1 Mathematical Model of the Glomerulus dq1 = K (P " P + ! ) ................(20.1) dx f 2 1 c Qi ! c = ! i .......................................................(20.4) qi # Q $ e %! Q & Q ' " e +! ln & i ' =1% K L i f ..............(20.5) Qi & 1%! ' !Qi & ' ( ) Qe =efflux through the efferent arterioles L=length of the filter !=" i /(P1 # P2) Cocurrent and Countercurrent Mechanism What is Cocurrent and Countercurrent Mechanism Why Study about it ? The human kidney use countercurrent exchange to remove water from urine so the body can retain water that was used to move the nitrogenous waste products. -
![L8-Urine Conc. [PDF]](https://docslib.b-cdn.net/cover/4402/l8-urine-conc-pdf-1384402.webp)
L8-Urine Conc. [PDF]
The loop of Henle is referred to as countercurrent multiplier and vasa recta as countercurrent exchange systems in concentrating and diluting urine. Explain what happens to osmolarity of tubular fluid in the various segments of the loop of Henle when concentrated urine is being produced. Explain the factors that determine the ability of loop of Henle to make a concentrated medullary gradient. Differentiate between water diuresis and osmotic diuresis. Appreciate clinical correlates of diabetes mellitus and diabetes insipidus. Fluid intake The total body water Antidiuretic hormone is controled by : Renal excretion of water Hyperosmolar medullary Changes in the osmolarity of tubular fluid : interstitium 1 2 3 Low osmolarity The osmolarity High osmolarity because of active decrease as it goes up because of the transport of Na+ and because of the reabsorbation of water co-transport of K+ and reabsorption of NaCl Cl- 4 5 Low osmolarity because of High osmolarity because of reabsorption of NaCl , also reabsorption of water in reabsorption of water in present of ADH , present of ADH reabsorption of urea Mechanisms responsible for creation of hyperosmolar medulla: Active Co- Facilitated diffusion transport : transport : diffusion : of : Na+ ions out of the Only of small thick portion of the K+ , Cl- and other amounts of water ascending limb of ions out of the thick from the medullary the loop of henle portion of the Of urea from the tubules into the into the medullary ascending limb of inner medullary medullary interstitium the loop of henle collecting -

Dinosaur Thermoregulation Were They “Warm Blooded”?
Department of Geological Sciences | Indiana University Dinosaurs and their relatives (c) 2015, P. David Polly Geology G114 Dinosaur thermoregulation Were they “warm blooded”? Thermogram of a lion Department of Geological Sciences | Indiana University Dinosaurs and their relatives (c) 2015, P. David Polly Geology G114 Body temperature in endotherms and ectotherms Thermogram of ostriches Thermogram of a snake wrapped around a human arm Thermogram of a python held by people Thermogram of a lion Department of Geological Sciences | Indiana University Dinosaurs and their relatives (c) 2015, P. David Polly Geology G114 Physiology, metabolism, and dinosaurs? Ornithopods Birds Sauropods Dromeosaurs Tyrannosaurus Ceratopsians Pachycephalosaurs Crocodilians Stegosaurs Ankylosaurs Lizards and snakes Lizards ? ? ? ? ? ? ? Department of Geological Sciences | Indiana University Dinosaurs and their relatives (c) 2015, P. David Polly Geology G114 “extant phylogenetic bracket” Using phylogenetic logic to reconstruct biology of extinct animals. Features observed in living animals can be traced back to common ancestor. This suggests that extinct clades that fall between are likely to have similar in features even if they cannot be observed directly in the fossils. Witmer, 1995 Department of Geological Sciences | Indiana University Dinosaurs and their relatives (c) 2015, P. David Polly Geology G114 Thermoregulation Maintaining body temperature within limited boundaries regardless of temperature of the surrounding environment. Endotherm vs. Ectotherm Endotherms use internal metabolic heat to regulate themselves, ectotherms use external sources of heat (and cool). Mammals and birds are endotherms, most other vertebrates are ectotherms. Homeotherm vs. Poikilotherm Homeotherms have a constant body temperature, poikilotherms have a variable body temperature. Mammals are homeotherms, but so are fish that live in water of constant body temperature. -
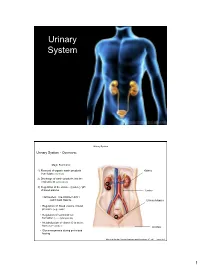
Urinary System
Urinary System Urinary System Urinary System - Overview: Major Functions: 1) Removal of organic waste products Kidney from fluids (excretion) 2) Discharge of waste products into the environment (elimination) 1 3) Regulation of the volume / [solute] / pH 3 of blood plasma Ureter HOWEVER, THE KIDNEY AIN’T JUST FOR PEE’IN… Urinary bladder • Regulation of blood volume / blood pressure (e.g., renin) • Regulation of red blood cell formation (i.e., erythropoietin) 2 • Metabolization of vitamin D to active form (Ca++ uptake) Urethra • Gluconeogenesis during prolonged fasting Marieb & Hoehn (Human Anatomy and Physiology, 8th ed.) – Figure 25.1 1 Urinary System Renal ptosis: Kidneys drop to lower position due Functional Anatomy - Kidney: to loss of perirenal fat Located in the superior lumbar “Bar of soap” region 12 cm x 6 cm x 3 cm 150 g / kidney Layers of Supportive Tissue: Renal fascia: Peritoneal cavity Outer layer of dense fibrous connective tissue; anchors kidney in place Perirenal fat capsule: Fatty mass surrounding kidney; cushions kidney against blows Fibrous capsule: Transparent capsule on kidney; prevents infection of kidney from local tissues Kidneys are located retroperitoneal Marieb & Hoehn (Human Anatomy and Physiology, 8th ed.) – Figure 25.2 Urinary System Functional Anatomy - Kidney: Pyelonephritis: Inflammation of the kidney Pyramids appear striped due to parallel arrangement of capillaries / collecting tubes Renal cortex Renal medulla Renal pyramids Renal papilla Renal columns Renal hilum Renal pelvis • Entrance for blood vessels -

AUTHOR Stevenson, R. D
DOCUMENT RESUME ED 195 429 SE 033 590 AUTHOR Stevenson, R.D. TITLE Animal Thermoregulation and the Operative Environmental (Equivalent) Temperature. Physical Processes in Terrestrial and Aquatic Ecosystems, Transport Process. INSTITUTION Washington Univ., Seattle. Center for Quantitative Science in Forestry, Fisheries and Wildlife. SPONS AGENCY National Science Foundation, Washington, D.C. PUB DATE Jun 79 GRANT NSF-GZ-2980: NSF-SED74-17696 NOTE 64p.: For related documents, see SE 033 581-597. EDFS PRICE MF01/PC03 Plus Postage. DESCRIPTORS *Biology: College Science: Computer Assisted Instruction: Computer Programs: Ecology: *Environmental Education: Heat: Higher Education: Instructional Materials: *Interdisciplinary Approach: Mathematical Applications: *Physiology: Science Education: Science Tnstruction: *Thermal Environment IDENTIFIERS *Thermoregulation ABSTRACT These materials were designed to be used by life sci,Ince students for instruction in the application of physical theory to ecosystem operation. Most modules contain computer programs which are built around a particular application of a physical process. Thermoregulation is defined as the ability of an organism to modify its body temperature. This module emphasizes animal thermoregulation from an ecological point of view and develops the concept of the operative environmental temperature, a technique which integrates the effects of all heat transfer processes into one number. Historical developments in the study of animal thermoregulation are reviewed and an attempt is made to illustrate how a quantitative knowledge of heat transfer processes can be used to clarify the adaptive significance of animal morphology, behavior, and physiology. A problem set illustrates and extends ideas developed in the text. (Author/CS1 *********************************************************************** Reproductions supplied by EDRS are the best that can be made from the original document. -

The External Environment
Chapter 2 The .xternal .n8ironment CHAPTER 2 THE EXTERNAL ENVIRONMENT Prepared for the Course Team by Patricia Ash 0.1 Introduction Before we can understand the phenotypic and genotypic adaptations of vertebrates for life in their environment we need to examine the physical characteristics of the environment. Vertebrates live both on land and in water so we need to study the thermal characteristics of each, and to understand the processes of heat exchange. We are especially interested in temperature because the rates of all metabolic processes are temperature dependent. As temperature decreases, enzyme activity usually decreases and so do the rates of most other chemical reactions. The highest temperature available for vertebrate metabolism is restricted by the risk of denaturation of proteins and disruption of membrane structure at temperatures greater than about 38°C. The body temperature of vertebrates represents a balance between heat gained either from metabolism or the environment and heat lost to the environment. Animal physiologists are therefore glucose particularly interested in thermal exchange. Metabolism CYTOSOL glycerol generates heat, and in endotherms this heat is used to maintain body temperature. Therefore, factors affecting metabolic rate alanine pyruvate have been much studied. Until recently most of this research was carried out in a laboratory, but techniques have been developed and refined that enable measurements of body acetyl CoA fatty acid temperature and metabolic rate to be carried out on animals in the field. aspartate 4C 6C MITOCHONDRIAL Nearly all vertebrates depend on oxygen for maintenance MATRIX of aerobic metabolism, and hence generation of ATP, in tissues. TCA cycle The highest yields of ATP per mole of substrate, are obtained glutamate from aerobic catabolism of fuel molecules. -

Characterization of the Heart Transcriptome of the White Shark (Carcharodon Carcharias) Richards Et Al
Photo courtesy of Michael Scholl - Save Our Seas Foundation Characterization of the heart transcriptome of the white shark (Carcharodon carcharias) Richards et al. Richards et al. BMC Genomics 2013, 14:697 http://www.biomedcentral.com/1471-2164/14/697 Richards et al. BMC Genomics 2013, 14:697 http://www.biomedcentral.com/1471-2164/14/697 RESEARCH ARTICLE Open Access Characterization of the heart transcriptome of the white shark (Carcharodon carcharias) Vincent P Richards1, Haruo Suzuki1,3, Michael J Stanhope1* and Mahmood S Shivji2* Abstract Background: The white shark (Carcharodon carcharias) is a globally distributed, apex predator possessing physical, physiological, and behavioral traits that have garnered it significant public attention. In addition to interest in the genetic basis of its form and function, as a representative of the oldest extant jawed vertebrate lineage, white sharks are also of conservation concern due to their small population size and threat from overfishing. Despite this, surprisingly little is known about the biology of white sharks, and genomic resources are unavailable. To address this deficit, we combined Roche-454 and Illumina sequencing technologies to characterize the first transciptome of any tissue for this species. Results: From white shark heart cDNA we generated 665,399 Roche 454 reads (median length 387-bp) that were assembled into 141,626 contigs (mean length 503-bp). We also generated 78,566,588 Illumina reads, which we aligned to the 454 contigs producing 105,014 454/Illumina consensus sequences. To these, we added 3,432 non-singleton 454 contigs. By comparing these sequences to the UniProtKB/Swiss-Prot database we were able to annotate 21,019 translated open reading frames (ORFs) of ≥ 20 amino acids.