The Association of Serum Biomarkers with Cardiac Health in Captive Gorillas
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

001 Emma the Bunny #006 Georgina the Hippo #011 Germaine
#001 #002 #003 #004 #005 Emma the bunny Alexandre the russian Piotr the polar bear Bridget the elephant Simon the sheep blue cat #006 #007 #008 #009 #010 Georgina the hippo Seamus the alpaca Austin the rhino Rufus the lion Richard the large white pig #011 #012 #013 #014 #015 Germaine the gorilla Winston the aardvark Penelope the bear Hank the dorset Fiona the panda down sheep #016 #017 #018 #019 #020 Juno the siamese cat Angharad the donkey Benedict the chimpanzee Samuel the koala Douglas the highland cow #021 #022 #023 #024 #025 Laurence the tiger Chardonnay the Claudia the Alice the zebra Audrey the nanny goat palomino pony saddleback pig #026 #027 #028 #029 #030 Clarence the bat Martin the tabby cat Sarah the friesian cow Timmy the jack russell Caitlin the giraffe #031 #032 #033 #034 #035 Esme the fox Blake the orangutan Siegfried the monkey Boris the red squirrel Hamlet the cheetah #036 #037 #038 #039 #040 Francis the hedgehog Jessie the raccoon Bradlee the grey squirrel Noah the zwartbles sheep Christophe the wolf #041 #042 #043 #044 #045 Sheila the kangaroo Andre the lemur Erica the Frank the armadillo Mae the snow leopard dromedary camel #046 #047 #048 #049 #050 Harriet the sloth Susan the badger Zack the skunk Donna the reindeer Logan the moose #051 #052 #053 #054 #055 Simone the suri alpaca Fred the herdwick sheep Eustice the beltex sheep Tobias the Seth the hebridean sheep wensleydale sheep #056 #057 #058 #059 #060 Tracy the racka sheep Xavier the gibbon Sid the giant anteater Caroline the platypus Harold the teeswater sheep -

Sleep and Nesting Behavior in Primates: a Review
Received: 29 July 2017 | Revised: 30 November 2017 | Accepted: 2 December 2017 DOI: 10.1002/ajpa.23373 REVIEW ARTICLE Sleep and nesting behavior in primates: A review Barbara Fruth1,2,3,4 | Nikki Tagg1 | Fiona Stewart2 1Centre for Research and Conservation/ KMDA, Antwerp, Belgium Abstract 2Faculty of Science/School of Natural Sleep is a universal behavior in vertebrate and invertebrate animals, suggesting it originated in the Sciences and Psychology, Liverpool John very first life forms. Given the vital function of sleep, sleeping patterns and sleep architecture fol- Moores University, United Kingdom low dynamic and adaptive processes reflecting trade-offs to different selective pressures. 3 Department of Developmental and Here, we review responses in sleep and sleep-related behavior to environmental constraints Comparative Psychology, Max-Planck- across primate species, focusing on the role of great ape nest building in hominid evolution. We Institute for Evolutionary Anthropology, Leipzig, Germany summarize and synthesize major hypotheses explaining the proximate and ultimate functions of 4Faculty of Biology/Department of great ape nest building across all species and subspecies; we draw on 46 original studies published Neurobiology, Ludwig Maximilians between 2000 and 2017. In addition, we integrate the most recent data brought together by University of Munich, Germany researchers from a complementary range of disciplines in the frame of the symposium “Burning the midnight oil” held at the 26th Congress of the International Primatological Society, Chicago, Correspondence Barbara Fruth, Liverpool John Moores August 2016, as well as some additional contributors, each of which is included as a “stand-alone” University, Faculty of Science/NSP, article in this “Primate Sleep” symposium set. -

Black Internationalism and African and Caribbean
BLACK INTERNATIONALISM AND AFRICAN AND CARIBBEAN INTELLECTUALS IN LONDON, 1919-1950 By MARC MATERA A Dissertation submitted to the Graduate School-New Brunswick Rutgers, the State University of New Jersey In partial fulfillment of the requirements For the degree of Doctor of Philosophy Graduate Program in History Written under the direction of Professor Bonnie G. Smith And approved by _______________________ _______________________ _______________________ _______________________ New Brunswick, New Jersey May 2008 ABSTRACT OF THE DISSERTATION Black Internationalism and African and Caribbean Intellectuals in London, 1919-1950 By MARC MATERA Dissertation Director: Bonnie G. Smith During the three decades between the end of World War I and 1950, African and West Indian scholars, professionals, university students, artists, and political activists in London forged new conceptions of community, reshaped public debates about the nature and goals of British colonialism, and prepared the way for a revolutionary and self-consciously modern African culture. Black intellectuals formed organizations that became homes away from home and centers of cultural mixture and intellectual debate, and launched publications that served as new means of voicing social commentary and political dissent. These black associations developed within an atmosphere characterized by a variety of internationalisms, including pan-ethnic movements, feminism, communism, and the socialist internationalism ascendant within the British Left after World War I. The intellectual and political context of London and the types of sociability that these groups fostered gave rise to a range of black internationalist activity and new regional imaginaries in the form of a West Indian Federation and a United West Africa that shaped the goals of anticolonialism before 1950. -

ANNEX 3 ICC-01/09-02/11-67-Anx3 21-04-2011 2/84 EO PT
ICC-01/09-02/11-67-Anx3 21-04-2011 1/84 EO PT No. ICC-01/09-02/11 21-4-11 ANNEX 3 ICC-01/09-02/11-67-Anx3 21-04-2011 2/84 EO PT A PROGRESS REPORT TO THE HON. ATTORNEY-GENERAL BY THE TEAM ON UPDATE OF POST ELECTION VIOLENCE RELATED CASES IN WESTERN, NYANZA, CENTRAL, RIFT-VALLEY, EASTERN, COAST AND NAIROBI PROVINCES MARCH, 2011 NAIROBI ICC-01/09-02/11-67-Anx3 21-04-2011 3/84 EO PT TABLE OF CONTENTS CHAPTER SUBJECT PAGE TRANSMITTAL LETTER IV 1. INTRODUCTION 1 2. GENDER BASED VIOLENCE CASES 7 3. WESTERN PROVINCE 24 3. RIFT VALLEY PROVINCE 30 4. NYANZA PROVINCE 47 5. COAST PROVINCE 62 6. NAIROBI PROVINCE 66 7. CENTRAL PROVINCE 69 8. STATISTICAL ANALYSIS 70 9. CONCLUSION 73 10. APPENDICES ICC-01/09-02/11-67-Anx3 21-04-2011 4/84 EO PT APPENDIX (NO.) LIST OF APPENDICES APP. 1A - Memo from CPP to Hon. Attorney General APP.1B - Memo from CPP to Hon. Attorney General APP.1C - Update on 2007 Post Election Violence offences As at 4th March, 2010 (police commissioner’s report) APP. 1D - Update by Taskforce on Gender Based Violence Cases (police commissioner’s report) APP. 2 - Memo to Solicitor- General from CPP APP. 3 - Letter from PCIO Western APP. 4 - Letter from PCIO Rift Valley APP.5 - Cases Pending Under Investigations in Rift Valley on special interest cases APP.6 - Cases where suspects are known in Rift Valley but have not been arrested APP.7 - Letter from PCIO Nyanza APP.8 - Letter from PCIO Coast APP.9 - Letter from PCIO Nairobi APP.10 - Correspondences from the team ICC-01/09-02/11-67-Anx3 21-04-2011 5/84 EO PT The Hon. -

High School Band Required List
2021-2022 High School Band Required List Table of Contents Click on the links below to jump to that list Band Group I (Required) Group II (Required) Group III (Required) BAND 2021‐2022 ISSMA REQUIRED LIST HIGH SCHOOL Perform as published unless otherwise specified. OOP = Permanently Out of Print * = New Additions or Changes this year Approximate times indicated for most selections GROUP I (REQUIRED) Aegean Festival Overture Makris / Bader E.C. Schirmer 10:00 Aerial Fantasy Michael A. Mogensen Barnhouse 10:30 Al Fresco Karel Husa Associated Music 12:13 American Faces David R. Holsinger TRN 6:00 American Hymnsong Suite (mvts. 1 & 3 or mvts. 1 & 4) Dwayne Milburn Kjos 3:11 / 1:45 / 1:50 American Overture For Band Joseph Willcox Jenkins Presser 115‐40040 5:00 American Salute Gould / Lang Belwin / Alfred 4:45 And The Mountains Rising Nowhere Joseph Schwantner European American Music 12:00 And The Multitude With One Voice Spoke James Hosay Curnow 8:00 Angels In The Architecture Frank Ticheli Manhattan Beach 15:00 Apotheosis Kathryn Salfelder Kon Brio 5:45 Apotheosis Of This Earth Karel Husa Associated Music 8:38 Archangel Raphael Leaves A House Of Tobias Masanori Taruya Foster Music 9:00 Armenian Dances Part I Alfred Reed Sam Fox / Plymouth 10:30 Armenian Dances Part II Alfred Reed Barnhouse 9:30 Army Of The Nile (March) Alford or Alford / Fennell Boosey & Hawkes 3:00 Aspen Jubilee Ron Nelson Boosey & Hawkes 10:00 Asphalt Cocktail John Mackey Osti Music 6:00 Austrian Overture Thomas Doss DeHaske 10:20 Away Day A. -

The Legal Apprentice
sja1-1_cv_sja1-1_cv 3/31/2014 2:40 PM Page 2 THE LEGAL APPRENTICE FOREWORD Dean Kathleen Voute’ MacDonald GOOD LEGAL WRITING IS SIMPLY GOOD WRITING: WORDS OF WELCOME Chief Judge Loretta A. Preska ARTICLES CONSTITUTIONAL BANS ON ALCOHOL ADS IN COLLEGE PRESS Shada Paula STARBUCKS’ BARRISTAS’ BATTLE: TIP POOL SHARING James Baek THE CASE OF THE CONNECTICUT CHIMP: STRICT LIABILITY FOR DOMESTICALLY KEPT WILD ANIMALS Jessica Fusco PROFESSIONAL PHOTOGRAPHER’S LIABILITY FOR REFUSAL TO WORK A SAME-SEX WEDDING Arturo Pena A LEGAL WRITING RESOURCE: THE BIBLE Professor Mary Noe Volume 1 Spring 2014 Number 1 sja1-1_cv_sja1-1_cv 3/31/2014 2:40 PM Page 3 The views expressed in the articles are to be attributed solely to their authors and not to St. John’s University or College of Professional Studies. Submissions to The Legal Apprentice or for additional copies contact [email protected]. ST. JOHN’S UNIVERSITY COLLEGE OF PROFESSIONAL STUDIES KATHLEEN VOUTE´ MACDONALD, ED.D. DEAN ELLEN TUFANO, PH.D. ASSOCIATE DEAN * * * JEFFREY GROSSMAN, J.D. CHAIRPERSON CRIMINAL JUSTICE, LEGAL STUDIES, HOMELAND SECURITY BERNARD HELLDORFER, J.D. DIRECTOR LEGAL STUDIES MARY NOE, J.D. ASSOCIATE PROFESSOR EDITOR JAMES CROFT, J.D. ASSISTANT PROFESSOR ASSISTANT EDITOR THE LEGAL APPRENTICE FACULTY ADVISORY BOARD JAMES CROFT, ASSISTANT PROFESSOR B.A., J. D. ALBANY UNIVERSITY ST. JOHN’S UNIVERSITY, SCHOOL OF LAW JUDITH CRAMER, ASSOCIATE PROFESSOR B.S., M.A., PH.D. UNIVERSITY OF HARTFORD THE UNION INSTITUTE AND UNIVERSITY, CINCINNATI, OH. SUSAN LUSHING, ASSOCIATE PROFESSOR B.S., J.D. CORNELL UNIVERSITY NEW YORK UNIVERSITY, SCHOOL OF LAW MARY NOE, ASSOCIATE PROFESSOR B.A., J.D. -

PROVISIONAL LIST.Pdf
S/N NAME YEAR OF CALL BRANCH PHONE NO EMAIL 1 JONATHAN FELIX ABA 2 SYLVESTER C. IFEAKOR ABA 3 NSIKAK UTANG IJIOMA ABA 4 ORAKWE OBIANUJU IFEYINWA ABA 5 OGUNJI CHIDOZIE KINGSLEY ABA 6 UCHENNA V. OBODOCHUKWU ABA 7 KEVIN CHUKWUDI NWUFO, SAN ABA 8 NWOGU IFIONU TAGBO ABA 9 ANIAWONWA NJIDEKA LINDA ABA 10 UKOH NDUDIM ISAAC ABA 11 EKENE RICHIE IREMEKA ABA 12 HIPPOLITUS U. UDENSI ABA 13 ABIGAIL C. AGBAI ABA 14 UKPAI OKORIE UKAIRO ABA 15 ONYINYECHI GIFT OGBODO ABA 16 EZINMA UKPAI UKAIRO ABA 17 GRACE UZOME UKEJE ABA 18 AJUGA JOHN ONWUKWE ABA 19 ONUCHUKWU CHARLES NSOBUNDU ABA 20 IREM ENYINNAYA OKERE ABA 21 ONYEKACHI OKWUOSA MUKOSOLU ABA 22 CHINYERE C. UMEOJIAKA ABA 23 OBIORA AKINWUMI OBIANWU, SAN ABA 24 NWAUGO VICTOR CHIMA ABA 25 NWABUIKWU K. MGBEMENA ABA 26 KANU FRANCIS ONYEBUCHI ABA 27 MARK ISRAEL CHIJIOKE ABA 28 EMEKA E. AGWULONU ABA 29 TREASURE E. N. UDO ABA 30 JULIET N. UDECHUKWU ABA 31 AWA CHUKWU IKECHUKWU ABA 32 CHIMUANYA V. OKWANDU ABA 33 CHIBUEZE OWUALAH ABA 34 AMANZE LINUS ALOMA ABA 35 CHINONSO ONONUJU ABA 36 MABEL OGONNAYA EZE ABA 37 BOB CHIEDOZIE OGU ABA 38 DANDY CHIMAOBI NWOKONNA ABA 39 JOHN IFEANYICHUKWU KALU ABA 40 UGOCHUKWU UKIWE ABA 41 FELIX EGBULE AGBARIRI, SAN ABA 42 OMENIHU CHINWEUBA ABA 43 IGNATIUS O. NWOKO ABA 44 ICHIE MATTHEW EKEOMA ABA 45 ICHIE CORDELIA CHINWENDU ABA 46 NNAMDI G. NWABEKE ABA 47 NNAOCHIE ADAOBI ANANSO ABA 48 OGOJIAKU RUFUS UMUNNA ABA 49 EPHRAIM CHINEDU DURU ABA 50 UGONWANYI S. AHAIWE ABA 51 EMMANUEL E. -

Pre-Sleep and Sleeping Platform Construction Behavior in Captive Orangutans (Pongo Spp
Original Article Folia Primatol 2015;86:187–202 Received: July 28, 2014 DOI: 10.1159/000381056 Accepted after revision: February 18, 2015 Published online: May 13, 2015 Pre-Sleep and Sleeping Platform Construction Behavior in Captive Orangutans (Pongo spp. ): Implications for Ape Health and Welfare a b–d David R. Samson Rob Shumaker a b Department of Evolutionary Anthropology, Duke University, Durham, N.C., Indianapolis c Zoo, VP of Conservation and Life Sciences, Indianapolis, Ind. , Indiana University, d Bloomington, Ind. , and Krasnow Institute at George Mason University, Fairfax, Va. , USA Key Words Orangutan · Sleep · Sleeping platform · Nest · Animal welfare · Ethology Abstract The nightly construction of a ‘nest’ or sleeping platform is a behavior that has been observed in every wild great ape population studied, yet in captivity, few analyses have been performed on sleep related behavior. Here, we report on such behavior in three female and two male captive orangutans (Pongo spp.), in a natural light setting, at the Indianapolis Zoo. Behavioral samples were generated, using infrared cameras for a total of 47 nights (136.25 h), in summer (n = 25) and winter (n = 22) periods. To characterize sleep behaviors, we used all-occurrence sampling to generate platform construction episodes (n = 217). Orangutans used a total of 2.4 (SD = 1.2) techniques and 7.5 (SD = 6.3) actions to construct a sleeping platform; they spent 10.1 min (SD – 9.9 min) making the platform and showed a 77% preference for ground (vs. elevated) sleep sites. Com- parisons between summer and winter platform construction showed winter start times (17:12 h) to be significantly earlier and longer in duration than summer start times (17:56 h). -

Pen International Writers in Prison Committee Caselist
PEN INTERNATIONAL WRITERS IN PRISON COMMITTEE CASELIST January-December 2013 PEN International Writers in Prison Committee 50/51 High Holborn London WC1V 6ER United Kingdom Tel: + 44 020 74050338 Fax: + 44 020 74050339 e-mail: [email protected] web site: www.pen-international.org PEN INTERNATIONAL CHARTER The P.E.N. Charter is based on resolutions passed at its International Congresses and may be summarised as follows: P.E.N. affirms that: 1. Literature knows no frontiers and must remain common currency among people in spite of political or international upheavals. 2. In all circumstances, and particularly in time of war, works of art, the patrimony of humanity at large, should be left untouched by national or political passion. 3. Members of P.E.N. should at all times use what influence they have in favour of good understanding and mutual respect between nations; they pledge themselves to do their utmost to dispel race, class and national hatreds, and to champion the ideal of one humanity living in peace in one world. 4. P.E.N. stands for the principle of unhampered transmission of thought within each nation and between all nations, and members pledge themselves to oppose any form of suppression of freedom of expression in the country and community to which they belong, as well as throughout the world wherever this is possible. P.E.N. declares for a free press and opposes arbitrary censorship in time of peace. It believes that the necessary advance of the world towards a more highly organized political and economic order renders a free criticism of governments, administrations and institutions imperative. -

201 ,QWHUQDWLRQDO 9Aluation Handbook ,QGXVWU\ Cost of Capital
201,QWHUQDWLRQDO9aluation Handbook ,QGXVWU\ Cost of Capital Market Results Through0DUFK 2015 Duff & Phelps &RPSDQ\/LVW 1RWH 7KLV GRFXPHQW SURYLGHV D OLVW RI WKH FRPSDQLHV XVHG WR SHUIRUP WKH DQDO\VHV SXEOLVKHG LQ WKH ,QWHUQDWLRQDO 9DOXDWLRQ +DQGERRN ̰ ,QGXVWU\ &RVW RI &DSLWDO GDWD WKURXJK 0DUFK 7KHLQIRUPDWLRQ KHUHLQ LV VSHFLILF WR WKH KDUGFRYHU ,QWHUQDWLRQDO 9DOXDWLRQ +DQGERRN ̰,QGXVWU\ &RVW RI &DSLWDO GDWD WKURXJK 0DUFK DQG LV QRW DSSOLFDEOH WR DQ\ RWKHU ERRN XSGDWH RU GRFXPHQW Cover image: Duff & Phelps Cover design: Tim Harms Copyright © 2016 by John Wiley & Sons, Inc. All rights reserved. Published by John Wiley & Sons, Inc., Hoboken, New Jersey. Published simultaneously in Canada. No part of this publication may be reproduced, stored in a retrieval system, or transmitted in any form or by any means, electronic, mechanical, photocopying, recording, scanning, or otherwise, except as permitted under Section 107 or 108 of the 1976 United States Copyright Act, without either the prior written permission of the Publisher, or authorization through payment of the appropriate per-copy fee to the Copyright Clearance Center, Inc., 222 Rosewood Drive, Danvers, MA 01923, (978) 750-8400, fax (978) 646-8600, or on the Web at www.copyright.com. Requests to the Publisher for permission should be addressed to the Permissions Department, John Wiley & Sons, Inc., 111 River Street, Hoboken, NJ 07030, (201) 748-6011, fax (201) 748-6008, or online at http://www.wiley.com/go/permissions. The foregoing does not preclude End-users from using the 2015 International Valuation Handbook ࣓ Industry Cost of Capital and data published therein in connection with their internal business operations. -
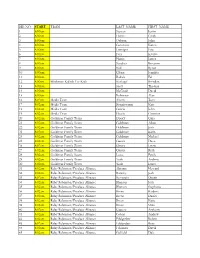
Step up List
BIB NO. START TEAM LAST_NAME FIRST_NAME 1 8:00am Stewart Justin 2 8:00am Harris Cindy 3 8:00am Osborn John 4 8:00am Geninatti Karen 5 8:00am Leninger Eric 6 8:00am Frey Kristin 7 8:00am Harris James 8 8:00am Sanchez Roxanne 9 8:00am Noll Byron 10 8:00am Glenn Jennifer 11 8:00am Badida Ed 12 8:00am Blackman Kallick For Kids Kinkopf Brendan 13 8:00am Scott Thomas 14 8:00am McGrath David 15 8:00am Robinson Alan 16 8:02am Media Team Ahern Tom 17 8:02am Media Team Bongiovanni Kate 18 8:02am Media Team Garcia John 19 8:02am Media Team Garcia Christine 20 8:02am Goldman Family Team Doocy Gina 21 8:02am Goldman Family Team Goldman Adam 22 8:02am Goldman Family Team Goldman Josh 23 8:02am Goldman Family Team Goldman Karla 24 8:02am Goldman Family Team Goldman Michael 25 8:02am Goldman Family Team Gussis Dave 26 8:02am Goldman Family Team Gussis Lizzie 27 8:02am Goldman Family Team Gussis Ruth 28 8:02am Goldman Family Team Lowe Emily 29 8:02am Goldman Family Team York Andrew 30 8:02am Goldman Family Team York Laura 31 8:02am Ruby Robinson/Produce Alliance Abrams Howard 32 8:02am Ruby Robinson/Produce Alliance Barasky Jodi 33 8:02am Ruby Robinson/Produce Alliance Bernstein Dustin 34 8:02am Ruby Robinson/Produce Alliance Blanton Josh 35 8:02am Ruby Robinson/Produce Alliance Blanton Stephanie 36 8:02am Ruby Robinson/Produce Alliance Breen Barbara 37 8:02am Ruby Robinson/Produce Alliance Breen Daniel 38 8:02am Ruby Robinson/Produce Alliance Breen Katie 39 8:02am Ruby Robinson/Produce Alliance Breen Mike 40 8:02am Ruby Robinson/Produce Alliance Capone -

NPRC) VIP List, 2009
Description of document: National Archives National Personnel Records Center (NPRC) VIP list, 2009 Requested date: December 2007 Released date: March 2008 Posted date: 04-January-2010 Source of document: National Personnel Records Center Military Personnel Records 9700 Page Avenue St. Louis, MO 63132-5100 Note: NPRC staff has compiled a list of prominent persons whose military records files they hold. They call this their VIP Listing. You can ask for a copy of any of these files simply by submitting a Freedom of Information Act request to the address above. The governmentattic.org web site (“the site”) is noncommercial and free to the public. The site and materials made available on the site, such as this file, are for reference only. The governmentattic.org web site and its principals have made every effort to make this information as complete and as accurate as possible, however, there may be mistakes and omissions, both typographical and in content. The governmentattic.org web site and its principals shall have neither liability nor responsibility to any person or entity with respect to any loss or damage caused, or alleged to have been caused, directly or indirectly, by the information provided on the governmentattic.org web site or in this file. The public records published on the site were obtained from government agencies using proper legal channels. Each document is identified as to the source. Any concerns about the contents of the site should be directed to the agency originating the document in question. GovernmentAttic.org is not responsible for the contents of documents published on the website.