Application of Forest Gap Model for Sal (Shorea Robusta) Forest Succession การประยกตุ ใช์ แบบจ้ าลองชํ ่องวางในป่ ่าสาหรํ ับการทดแทนของป่าซาล ์
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Litsea Monopetala Lauraceae
Litsea monopetala (Roxb.) Persoon Lauraceae LOCAL NAMES Burmese (ondon laukya); Hindi (maidalagadil,singran,bastuva,rapamba,muga,meda,kalmara,kavkawa); Indonesian (huru manuk); Malay (bangang); Nepali (kutmiro,soalu,ratmanti,kutmero); Thai (kathang) BOTANIC DESCRIPTION Litsea monopetala is a small tree up to 18 m tall, bole straight to crooked, up to 60 cm in diameter, bark surface longitudinally fissured, dark greyish, inner bark brown mottled. Leaves alternate, 4.5-17 cm x 2.5-10 cm, blunt to acute, glabrous above, sparsely hairy below, midrib sunken above, with 6-13 pairs of secondary veins which are sunken above, tertiary venation scalariform, distinct below, petiole 1-2.5 cm long. Flowers in peduncled umbellules in short racemes, with (4-)6 tepals and 9- 12 stamens. Fruit oblong to ellipsoid, 0.7-1.2 cm long, seated on a small flat perianth cup. BIOLOGY Flower appears in March-April and fruit ripen in July-August. The fruits of L. monopetala are thought to be dispersed by bats. Agroforestry Database 4.0 (Orwa et al.2009) Page 1 of 5 Litsea monopetala (Roxb.) Persoon Lauraceae ECOLOGY L. monopetala is locally common in mixed lowland and montane evergreen or semi-deciduous forest. BIOPHYSICAL LIMITS Altitude: Up to 1500 m. DOCUMENTED SPECIES DISTRIBUTION Native: China, India, Indonesia, Myanmar, Thailand Exotic: Native range Exotic range The map above shows countries where the species has been planted. It does neither suggest that the species can be planted in every ecological zone within that country, nor that the species can not be planted in other countries than those depicted. -

Insect Fauna Associated with Muga-Ecosystem in North East India and Ambrosia Beetle
Invasive Ambrosia Beetle Conference The Situation in California August 12 – 14, 2012 Meeting sponsored by: The Hofshi Foundation University of California, Riverside UC Center for Invasive Pest Research The Huntington Botanical Gardens The Los Angeles Arboretum Invasive Ambrosia Beetle Conference The Situation in California August 12 – 14, 2012 Session 6 Invasion Epidemiology INSECT FAUNA ASSOCIATED WITH MUGA-ECOSYSTEM IN NORTH EAST INDIA AND AMBROSIA BEETLE DR. RAJESH KUMAR CENTRAL MUGA, ERI RESEARCH & TRAINING INSTITUTE CENTRAL SILK BOARD MINISTRY OF TEXTILES, GOVT. OF INDIA Lahdoigarh – 785 700 Jorhat (ASSAM) EXPLORATION Central Muga Eri Sikkim Arunachal Pradesh Research and Training Institute, Meghalaya Jorhat - ASSAM 1200cm rainfall at Humidity : 85-100% some Temp: 25-32ºC places ASSAM Nagaland Tripura Mizoram Manipur FOUR TYPES OF SILLKWORMS IN NORTH EAST INDIA SILKWORMS AND HOST PLANTS Som, Persea bombycina Soalu , Litsea monopetala Antheraea assamensis Helfer Digloti, L. salicifolia Mejankri, L. citrata Antheraea pernyi Samia ricini Boisd. Castor, Ricinus communis) Tapioca , Manihot esculenta Oak, Quercus serrata, Kesseru, Heteropanax Q. dealbata fragrans Payam, Evodia fraxinifolia, Bombyx mori Linn. Barpat, Ailanthus grandis, A. excelsa A. altissima Mulberry, Morus spp. III instar I instar II instar IV instar V instar 22/03/2012 Rearing in the natural condition Collection of silkworms Collection of cocoons Traditionally called “Jali” (dried branches of Persea bombycina Kost) Harvested Cocoons Muga Silksworm Cocoons Muga Silksworm Femle moths tied on small branch of som plants for laying eggs Muga Silk – A. assamensis Eri Silk – Samia ricini PRELIMINARY STUDY Conducted at Farms of the Institute at Lahdoigarh, Jorhat and at farmers field of Sivasagar and Golaghat District of Assam on two host plants of Muga Silkworm (Antheraea assamensis Helfer) Som (Persea bombycina Kost ) and Soalu (Litsea monopetala). -

Ethnopharmacological Properties and Medicinal Uses of Litsea Cubeba
plants Review Ethnopharmacological Properties and Medicinal Uses of Litsea cubeba Madhu Kamle 1, Dipendra K. Mahato 2, Kyung Eun Lee 3, Vivek K. Bajpai 4, Padam Raj Gajurel 1, Kang Sang Gu 3,5,* and Pradeep Kumar 1,* 1 Department of Forestry, North Eastern Regional Institute of Science and Technology, Nirjuli 791109, India; [email protected] (M.K.); [email protected] (P.R.G.) 2 School of Exercise and Nutrition Sciences, Deakin University, Burwood, VIC 3125, Australia; [email protected] 3 Molecular Genetics Lab, Department of Biotechnology, Yeungnam University, Gyeongsan, Gyeongbuk 38541, Korea; [email protected] 4 Department of Energy and Material Engineering, Dongguk University-Seoul, Seoul 04620, Korea; [email protected] 5 Stemforce, 302 Institute of Industrial Technology, Yeungnam University, Gyeongsan, Gyeongbuk 38541, Korea; [email protected] * Correspondence: [email protected] (K.S.G.); [email protected] (P.K.) Received: 8 May 2019; Accepted: 30 May 2019; Published: 1 June 2019 Abstract: The genus Litsea is predominant in tropical and subtropical regions of India, China, Taiwan, and Japan. The plant possesses medicinal properties and has been traditionally used for curing various gastro-intestinal ailments (e.g., diarrhea, stomachache, indigestion, and gastroenteritis) along with diabetes, edema, cold, arthritis, asthma, and traumatic injury. Besides its medicinal properties, Litsea is known for its essential oil, which has protective action against several bacteria, possesses antioxidant and antiparasitic properties, exerts acute and genetic toxicity as well as cytotoxicity, and can even prevent several cancers. Here we summarize the ethnopharmacological properties, essentials oil, medicinal uses, and health benefits of an indigenous plant of northeast India, emphasizing the profound research to uplift the core and immense potential present in the conventional medicine of the country. -

WRA Species Report
Family: Salicaceae Taxon: Flacourtia indica Synonym: Flacourtia ramontchi L’Hér. Common Name: batoko plum Flacourtia sepiaria Roxb. governor's plum Gmelina indica Burm. f. (basionym) Indian plum Madagascar plum ramontchi Questionaire : current 20090513 Assessor: Chuck Chimera Designation: H(HPWRA) Status: Assessor Approved Data Entry Person: Chuck Chimera WRA Score 12 101 Is the species highly domesticated? y=-3, n=0 n 102 Has the species become naturalized where grown? y=1, n=-1 103 Does the species have weedy races? y=1, n=-1 201 Species suited to tropical or subtropical climate(s) - If island is primarily wet habitat, then (0-low; 1-intermediate; 2- High substitute "wet tropical" for "tropical or subtropical" high) (See Appendix 2) 202 Quality of climate match data (0-low; 1-intermediate; 2- High high) (See Appendix 2) 203 Broad climate suitability (environmental versatility) y=1, n=0 y 204 Native or naturalized in regions with tropical or subtropical climates y=1, n=0 y 205 Does the species have a history of repeated introductions outside its natural range? y=-2, ?=-1, n=0 y 301 Naturalized beyond native range y = 1*multiplier (see y Appendix 2), n= question 205 302 Garden/amenity/disturbance weed n=0, y = 1*multiplier (see Appendix 2) 303 Agricultural/forestry/horticultural weed n=0, y = 2*multiplier (see n Appendix 2) 304 Environmental weed n=0, y = 2*multiplier (see y Appendix 2) 305 Congeneric weed n=0, y = 1*multiplier (see y Appendix 2) 401 Produces spines, thorns or burrs y=1, n=0 y 402 Allelopathic y=1, n=0 403 Parasitic y=1, -

Effect of Different Environmental Conditions on Germination of Two Major Species of Litsea (Lauracae) of Uttarakhand Himalaya
Octa Journal of Environmental Research Apr. – Jun., 2015 International Peer-Reviewed Journal ISSN 2321 3655 Oct. Jour. Env. Res. Vol. 3(2): 144-152 Available online http://www.sciencebeingjournal.com Research Article EFFECT OF DIFFERENT ENVIRONMENTAL CONDITIONS ON GERMINATION OF TWO MAJOR SPECIES OF LITSEA (LAURACAE) OF UTTARAKHAND HIMALAYA Kavita Tariyal Department of Applied Sciences and Humanities, THDC Institute of Hydropower Engineering and Technology, Bhagirthipuram, Tehri Garhwal- 249001, Uttarakhand, India *Corresponding author’s Email: [email protected] Received: 06th June 2015 Revised: 22nd June 2015 Accepted: 25th June 2015 Abstract: Present investigation covers monitoring on seed germination (under the influence of different growth regulators) on two important tree species of the region, viz- Litsea elongata, and Litsea monopetala. This analysis was considered necessary since the beneficial effect of pre-soaking treatment of seeds with growth regulators and other substances have been reported in the literature repeatedly. It is now generally agreed that such a treatment leads to an increase in tissue hydration, respiratory activity, redistribution of nutrient reserves, and enhancement of the seedling growth, thus it is bound to help us in knowing the causes of the dormancy of seeds in forest tree species on the one hand and seems to be the potential approach to keep pace with new concepts and developments to meet the increasing demands of the country especially through the release of new seedling and the samplings growth -

3. LITSEA Lamarck, Encycl. 3: 574. 1792, Nom. Cons
Flora of China 7: 118–141. 2008. 3. LITSEA Lamarck, Encycl. 3: 574. 1792, nom. cons. 木姜子属 mu jiang zi shu Huang Puhua (黄普华 Huang Pu-hwa), Li Jie (李捷), Li Xiwen (李锡文 Li Hsi-wen); Henk van der Werff Hexanthus Loureiro; Iozoste Nees; Malapoënna Adanson; Pseudolitsea Yen C. Yang; Tetranthera Jacquin. Trees or shrubs, evergreen or deciduous, dioecious. Leaves alternate, rarely opposite or verticillate, pinninerved. Umbels, or umbellate cymes or panicles, solitary or clustered in leaf axils; involucral bracts 4–6, decussate, persistent at flowering. Perianth tube long or short, segments usually 6, in 2 whorls of 3 each, equal or unequal, rarely lacking or 8. Flowers unisexual. Male flowers: fer- tile stamens 9 or 12, rarely more, in 3 or 4 whorls of 3 each; filaments of 1st and 2nd whorls usually eglandular, of 3rd and 4th 2- glandular at base; anthers all introrse, 4-celled, cells opening by lids; rudimentary pistil present or lacking. Female flowers: stam- inodes as many as stamens of male flowers; ovary superior; style conspicuous. Fruit seated on perianth tube; perianth tube ± enlarged, shallowly discoid or deeply cup-shaped or unaltered at fruit. About 200 species: mainly in tropical and subtropical Asia, a few species in Australia and from North America to subtropical South America; 74 species (47 endemic) in China. Litsea mishmiensis J. D. Hooker (Fl. Brit. India 5: 161. 1886; see FRPS 31: 336. 1982) was described from the border region between NE India and SE Xizang. Tetranthera floribunda Champion ex Bentham (Hooker’s J. Bot. Kew Gard. Misc. 5: 199. -
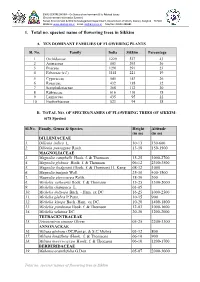
1. Total No. Species/ Name of Flowering Trees in Sikkim
ENVIS CENTRE SIKKIM – On Status of environment & Its Related Issues (Environmental Information System) Forest, Environment & Wildlife Management Department, Government of Sikkim, Deorali, Gangtok – 737102 Website: www.sikenvis.nic.in Email: [email protected] Tele/Fax: 03592-280381 1. Total no. species/ name of flowering trees in Sikkim A. TEN DOMINANT FAMILIES OF FLOWERING PLANTS Sl. No. Family India Sikkim Percentage 1 Orchidaceae 1229 527 43 2 Asteraceae 803 293 36 3 Poaceae 1291 291 23 4 Fabaceae (s.l.) 1141 221 19 5 Cyperaceae 545 143 26 6 Rosaceae 432 138 32 7 Scrophulariaceae 368 112 30 8 Rubiaceae 616 110 18 9 Lamiaceae 435 95 22 10 Euphorbiaceae 523 94 18 B. TOTAL NO. OF SPECIES/NAMES OF FLOWERING TREES OF SIKKIM: (678 Species) Sl.No. Family, Genus & Species Height Altitude (in m) (in m) DILLENIACEAE 1. Dillenia indica L. 10-13 150-600 2. Dillenia pentagyna Roxb. 13-18 150-1500 MAGNOLIACEAE 3. Magnolia campbelli Hook. f. & Thomson 15-25 1000-2700 4. Magnolia globosa Hook. f. & Thomson 06-12 2300-3500 5. Magnolia hodgsonii (Hook. f. & Thomson) H. Keng 08-12 1000 6. Magnolia insignis Wall. 25-30 400-1500 7. Magnolia pterocarpa Roxb. 18-30 500 8. Michelia cathcartii Hook. f. & Thomson 15-25 1500-2000 9. Michelia champaca L. 03-05 10. Michelia doltsopa Buch. - Ham. ex DC. 16-25 1000-2500 11. Michelia glabra P.Pann. 10-15 900 12. Michelia kisopa Buch.-Ham. ex DC. 10-20 1400-1800 13. Michelia punduana Hook. f. & Thomson 12-03 1000-1600 14. Michelia velutina DC. -

9 Importance of Trees Outside Forest (Tof)
DOI: https://doi.org/10.3329/jbcbm.v6i1.51326 J. biodivers. conserv. bioresour. manag. 6(1), 2020 IMPORTANCE OF TREES OUTSIDE FOREST (TOF) FOR IMMEDIATE EARTHQUAKE RESPONSE Ghimire, P., P. Paudel1, B. Bhatta, P. Gautam1 and N.Devkota2 Faculty of Forestry, Agriculture and Forestry University, Hetauda, Nepal; 1Practical Solution Consultancy Nepal, Pvt. Ltd., Kathmandu, Nepal; 2Faculty of Agriculture, Agriculture and Forestry University, Chitwan, Nepal Abstract A study was carried out in Thulosirubari village of Chautara Sangachowkgadi municipality of Sindhupalchok district, Nepal to analyse the importance of TOF for immediate earthquake response and recovery. More than 80% of the households have trees in their own land with species such as Chilaune (Schima wallichii), Kutmiro (Litsea monopetala), Khote Salla (Pinus roxburghii) and Bamboo are common. Among surveyed households (134), more than 75% respondents reported that they used Schima wallichii for pillar (Khaba) to make temporary houses, whereas bamboo was used for all kind of poles (Dada/Vata/Balo) in more than 80% of temporary houses. Both Bamboo and Schima wallichii were either extracted from their own land or from neighbour's land, i.e. TOF which were easily available and collected as required. We could not draw any statistical conclusion in choice of species to construct temporary houses by wealth categories. However, we have observed that people who have sufficient and varieties of trees in own land, have selected strong and hard wood for permanent house construction whereas those who do not have choice, are limited to use any of the available tree species. People were highly aware about importance of trees to cope with disaster; however selection of species was still debatable because of limited knowledge and researches. -

Conservation Issues of Litsea Monopetala (Roxb.) Persoon (Lauracae) in Pakistan
Pak. J. Bot., 52(5): 1697-1706, 2020. DOI: http://dx.doi.org/10.30848/PJB2020-5(45) CONSERVATION ISSUES OF LITSEA MONOPETALA (ROXB.) PERSOON (LAURACAE) IN PAKISTAN FAZAL AMIN, HAIDAR ALI AND HASSAN SHER Center for Plant Sciences and Biodiversity, University of Swat *Corresponding author's email: [email protected] Abstract Economically important especially medicinal plants are facing threats of habitat alteration, habitat degradation, unplanned urbanization, over exploitation and unsustainable use practices and these threats are mainly responsible for the current species extirpation. Litsea monopetala is economically important medicinal plant. In Pakistan it was previously reported from Salt Range only, since, no specific locality was mentioned in the literature, therefore, in the present study L. monopetala could not be located in Salt Range in spite of our repeated efforts. However, after four years of continuous field studies during 2015 to 2018, three new localities were discovered i.e. Aba Chena Najigram (district Swat), Shoprang and Char Tambo (district Buner). A total of 218 mature trees were found in 3 localities in 2015, 214 mature trees were found in 3 localities in 2016, 192 mature trees were found in 3 localities in 2017, whereas, 179 mature trees were found in 1 locality i.e. Shoprang in 2018. Whereas, no individuals were found in Aba Chena Najigram and Char Tamboo during the year 2018. During the four years study in these 3 localities 1.84%, 10.28% and 6.77% decrease has been observed in its total population size, respectively. The main threats faced by the taxon were its habitat alteration, agricultural land extension, unsustainable lopping and bark pealing. -

The Hemiptera-Sternorrhyncha (Insecta) of Hong Kong, China—An Annotated Inventory Citing Voucher Specimens and Published Records
Zootaxa 2847: 1–122 (2011) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Monograph ZOOTAXA Copyright © 2011 · Magnolia Press ISSN 1175-5334 (online edition) ZOOTAXA 2847 The Hemiptera-Sternorrhyncha (Insecta) of Hong Kong, China—an annotated inventory citing voucher specimens and published records JON H. MARTIN1 & CLIVE S.K. LAU2 1Corresponding author, Department of Entomology, Natural History Museum, Cromwell Road, London SW7 5BD, U.K., e-mail [email protected] 2 Agriculture, Fisheries and Conservation Department, Cheung Sha Wan Road Government Offices, 303 Cheung Sha Wan Road, Kowloon, Hong Kong, e-mail [email protected] Magnolia Press Auckland, New Zealand Accepted by C. Hodgson: 17 Jan 2011; published: 29 Apr. 2011 JON H. MARTIN & CLIVE S.K. LAU The Hemiptera-Sternorrhyncha (Insecta) of Hong Kong, China—an annotated inventory citing voucher specimens and published records (Zootaxa 2847) 122 pp.; 30 cm. 29 Apr. 2011 ISBN 978-1-86977-705-0 (paperback) ISBN 978-1-86977-706-7 (Online edition) FIRST PUBLISHED IN 2011 BY Magnolia Press P.O. Box 41-383 Auckland 1346 New Zealand e-mail: [email protected] http://www.mapress.com/zootaxa/ © 2011 Magnolia Press All rights reserved. No part of this publication may be reproduced, stored, transmitted or disseminated, in any form, or by any means, without prior written permission from the publisher, to whom all requests to reproduce copyright material should be directed in writing. This authorization does not extend to any other kind of copying, by any means, in any form, and for any purpose other than private research use. -

A Study on Litsea Monopetala for Evaluation of Pharmacological Activities Md
DOI: 10.21276/sjmps Saudi Journal of Medical and Pharmaceutical Sciences ISSN 2413-4929 (Print) Scholars Middle East Publishers ISSN 2413-4910 (Online) Dubai, United Arab Emirates Website: http://scholarsmepub.com/ Original Research Article A Study on Litsea monopetala for Evaluation of Pharmacological Activities Md. Shohel Hossain1, Mohammad Tarek Aziz2, Khondoker Md. Akter-uz-zaman Sumon1, Md. Ibrahim3*, Abida Sultana4, Nane Gopal Sarker5, Palash Das3, Md. Masudur Rahman3, Md. Jahirul Islam3, Mohammad Hasem Babu3 1Department of Pharmacy, Manarat International University, Bangladesh 2Department of Pharmacy, State University of Bangladesh, Bangladesh 3Department of Pharmacy, Atish Dipankar University of Science and Technology, Bangladesh 4Department of Applied Chemistry and Chemical Engeering, Noakhali Science and Technology University, Bangladesh 5Department of Pharmacy, University of Development Alternative, Bangladesh *Corresponding Author: Md. Ibrahim Email: [email protected] Abstract: The aim of this study was to evaluate the cytotoxicity, thrombolytic, anthelmintic and antioxidant activity of mehanolic extract of Litsea monopetala (Family: Lauraceae) leaves in laboratory using in vitro methods. Cytotoxicity test was done by brine shrimp lethality bioassay where the extract concentration was 400, 200, 100, 50, 25, 12.5, 6.25, 3.125, 1.5625, 0.78125 (µg/ml). In vitro thrombolytic activity was performed by clot lysis method using extract concentration 2.5, 5, 10 and 20 (mg/mL) in saline water. Anthelmintic activity test was done by using adult earthworms where 10, 20, 40, 60, 80 (mg/ml) extract concentration were used. Finally antioxidant activity was determined by total phenolic content determination using Folin-Ciocalteu reagent. The crude extract of Litsea monopetala showed cytotoxic activity against brine shrimp nauplii and LC50 value was 41.05(µg/ml) and the investigated thrombolytic activity in our research was 9.52,9.49, 13.64 and 17.50 % respectively as % of clot lysis. -
![Diversity of Litsea Lamarck [Lauraceae] in Terai and Duars Regions of West Bengal, India](https://docslib.b-cdn.net/cover/3565/diversity-of-litsea-lamarck-lauraceae-in-terai-and-duars-regions-of-west-bengal-india-3173565.webp)
Diversity of Litsea Lamarck [Lauraceae] in Terai and Duars Regions of West Bengal, India
Pleione 8(1): 68 - 78. 2014. ISSN: 0973-9467 © East Himalayan Society for Spermatophyte Taxonomy Diversity of Litsea Lamarck [Lauraceae] in Terai and Duars regions of West Bengal, India Dibakar Choudhury1, Rajib Biswas1, Palash Mandal2 and A. P. Das1* 1Taxonomy and Environmental Biology Laboratory, Department of Botany, University of North Bengal, West Bengal, India 2Plant Physiology and Pharmacognosy Research Laboratory, Department of Botany, University of North Bengal, West Bengal, India * Corresponding author, E- mails: [email protected] [Received 23.03.2014; Revised 12.05.2014; Accepted 29.05.2014; Published 30.06.2014] Abstract There are about 622 species in the genus Litsea Lamarck (Lauraceae), distributed mainly in Australia, New Zealand, North & South America and Asia. Among those, 45 species of Litsea were reported from India. No comprehensive report is available on this genus from Terai and Duars regions, which are located at the foot of the Eastern Himalaya and located within the Himalaya Hotspot. Present paper reported the occurrence of nine species of Litsea from Terai and Duars belt along with their local names, salient features, exsiccatus, status, flowering and fruiting periods and geographic distribution. Collection of L. assamica Hooker f. from the Jaldapara National Park in Duars forms the new record for the state flora of West Bengal. Key words: Genus Litsea, Terai, Duars, West Bengal, Litsea assamica, new record INTRODUCTION The Terai and Duars region politically constitute the plains of Darjeeling and whole of Jaipaiguri districts of West Bengal. Northern part of West Bengal touching the feat of Eastern Himalaya is generally referred as Terai (25º 57' to 26° 36' N, Latitude and 89° 54' to 88° 47' longitude) and Duars (located between 26° 16' to 27° 0' N latitude and 88° 4' to 89° 53' E longitude) (Ghosh 2006; Das et al 2010; Roy et al 2009).