Archeological Evidence of Large Northern Bluefin Tuna
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Gary's Charts
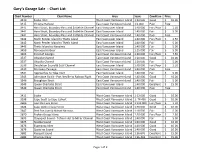
Gary’s Garage Sale - Chart List Chart Number Chart Name Area Scale Condition Price 3410 Sooke Inlet West Coast Vancouver Island 1:20 000 Good $ 10.00 3415 Victoria Harbour East Coast Vancouver Island 1:6 000 Poor Free 3441 Haro Strait, Boundary Pass and Sattelite Channel East Vancouver Island 1:40 000 Fair/Poor $ 2.50 3441 Haro Strait, Boundary Pass and Sattelite Channel East Vancouver Island 1:40 000 Fair $ 5.00 3441 Haro Strait, Boundary Pass and Sattelite Channel East Coast Vancouver Island 1:40 000 Poor Free 3442 North Pender Island to Thetis Island East Vancouver Island 1:40 000 Fair/Poor $ 2.50 3442 North Pender Island to Thetis Island East Vancouver Island 1:40 000 Fair $ 5.00 3443 Thetis Island to Nanaimo East Vancouver Island 1:40 000 Fair $ 5.00 3459 Nanoose Harbour East Vancouver Island 1:15 000 Fair $ 5.00 3463 Strait of Georgia East Coast Vancouver Island 1:40 000 Fair/Poor $ 7.50 3537 Okisollo Channel East Coast Vancouver Island 1:20 000 Good $ 10.00 3537 Okisollo Channel East Coast Vancouver Island 1:20 000 Fair $ 5.00 3538 Desolation Sound & Sutil Channel East Vancouver Island 1:40 000 Fair/Poor $ 2.50 3539 Discovery Passage East Coast Vancouver Island 1:40 000 Poor Free 3541 Approaches to Toba Inlet East Vancouver Island 1:40 000 Fair $ 5.00 3545 Johnstone Strait - Port Neville to Robson Bight East Coast Vancouver Island 1:40 000 Good $ 10.00 3546 Broughton Strait East Coast Vancouver Island 1:40 000 Fair $ 5.00 3549 Queen Charlotte Strait East Vancouver Island 1:40 000 Excellent $ 15.00 3549 Queen Charlotte Strait East -

Yellowfin Tuna (Thunnus Albacares) and Albacore (Thunnus Alalunga) (Perciformes: Scombrinae) in the Atlantic, Indian and Eastern Pacific Oceans
SCRS/2009/024 Collect. Vol. Sci. Pap. ICCAT, 65(2): 717-724 (2010) LENGTH-WEIGHT RELATIONSHIPS FOR BIGEYE TUNA (THUNNUS OBESUS), YELLOWFIN TUNA (THUNNUS ALBACARES) AND ALBACORE (THUNNUS ALALUNGA) (PERCIFORMES: SCOMBRINAE) IN THE ATLANTIC, INDIAN AND EASTERN PACIFIC OCEANS Guoping Zhu, Liuxiong Xu*,Yingqi Zhou, Liming Song and Xiaojie Dai SUMMARY The length-weight relationships (LWRs) for bigeye tuna (Thunnus obesus), yellowfin tuna (Thunnus albacares) and albacore (Thunnus alalunga) collected in the Atlantic, Indian and Eastern Pacific Oceans are presented. Significant differences can be found from the fork length distributions and the LWRs of the above 3 tuna species and the relations of dressed weight and round weight of bigeye and yellowfin tunas collected from the Atlantic, Indian and Eastern Pacific Oceans. The growth type of bigeye and yellowfin tuna in the Eastern Pacific Ocean showed positive allometries, whereas bigeye and yellowfin tuna from the Atlantic and Indian Oceans and albacore from the Atlantic, Indian and Eastern Pacific Oceans indicated negative allometries. RÉSUMÉ Le présent document décrit les relations longueur-poids (LWR) du thon obèse (Thunnus obesus), de l’albacore (Thunnus albacares) et du germon (Thunnus alalunga) prélevés dans les océans Atlantique, Indien et Pacifique de l’Est. Des différences considérables peuvent être trouvées dans les distributions longueur-fourche et les relations longueur-poids des trois espèces thonières susmentionnées et dans les relations du poids manipulé et du poids vif du thon obèse et de l’albacore collectés dans les océans Atlantique, Indien et Pacifique Est. Le type de croissance du thon obèse et de l’albacore dans l’océan Pacifique Est a révélé des allométries positives, tandis que le thon obèse et l’albacore des océans Atlantique et Indien et le germon des océans Atlantique, Indien et Pacifique Est ont fait apparaître des allométries négatives. -

Atlantic Bluefin Tuna (Thunnus Thynnus) Population Dynamics
Environ. Sci. Technol. 2009, 43, 8522–8527 nations of the International Commission for the Conservation Atlantic Bluefin Tuna (Thunnus of Atlantic Tunas (ICCAT) currently manage ABFT fisheries thynnus) Population Dynamics assuming two units (a western stock spawning in the Gulf of Mexico, and an eastern stock which spawns in the Delineated by Organochlorine Mediterranean Sea) ostensibly separated by the 45° W meridian with little intermixing between stocks. However, Tracers tagging studies indicate that bluefin tuna undergo extensive and complex migrations, including trans-Atlantic migrations, ,† and that stock mixing could be as high as 30% (2-4). Extensive REBECCA M. DICKHUT,* - ASHOK D. DESHPANDE,‡ mixing of eastern and western stocks (35 57% bluefin tuna ALESSANDRA CINCINELLI,§ of eastern origin) within the U.S. Mid Atlantic Bight was also 18 MICHELE A. COCHRAN,† reported recently based on otolith δ O values (5). The SIMONETTA CORSOLINI,| uncertainty of stock structures due to mixing makes it difficult RICHARD W. BRILL,† DAVID H. SECOR,⊥ for fisheries managers to assess the effectiveness of rebuilding AND JOHN E. GRAVES† efforts for the dwindling western Atlantic spawning stock of Virginia Institute of Marine Science, Gloucester Point, bluefin tuna. Understanding ABFT spatial distributions and Virginia 23062, National Marine Fisheries Service, dynamics are vital for robust population assessments and Highlands, New Jersey 07732, Department of Chemistry, the design of effective management strategies, and there is University of Florence, 50019 -

A Preliminary Study on the Stomach Content of Southern Bluefin Tuna Thunnus Maccoyii Caught by Taiwanese Longliner in the Central Indian Ocean
CCSBT-ESC/0509/35 A preliminary study on the stomach content of southern bluefin tuna Thunnus maccoyii caught by Taiwanese longliner in the central Indian Ocean Kwang-Ming Liu1, Wei-Ke Chen2, Shoou-Jeng Joung2, and Sui-Kai Chang3 1. Institute of Marine Resource Management, National Taiwan Ocean University, Keelung, Taiwan. 2. Department of Environmental Biology and Fisheries Science, National Taiwan Ocean University, Keelung, Taiwan. 3. Fisheries Agency, Council of Agriculture, Taipei, Taiwan. Abstract The stomach contents of 63 southern bluefin tuna captured by Taiwanese longliners in central Indian Ocean in August 2004 were examined. The size of tunas ranged from 84-187 cm FL (12-115 kg GG). The length and weight frequency distributions indicated that most specimens were in the range of 100-130 cm FL with a body weight between 10 and 30 kg for both sexes. The sexes- combined relationship between dressed weight and fork length can be described by W = 6.975× 10-6× FL3.1765 (n=56, r2=0.967, p < 0.05). The subjective index of fullness of specimens was estimated as: 1 = empty (38.6%), 2 = <half full (47.37%), 3 = half full (3.51%), 4 = >half full (5.26%), and 5 = full (5.26%). For the stomachs with prey items, almost all the preys are pisces and the proportion of each prey groups are fishes (95.6%), cephalopods (2.05%), and crustaceans (0.02%). In total, 6 prey taxa were identified – 4 species of fish, 1 unidentified pisces, 1 unidentified crustacean, and 1 unidentified squid. The 4 fish species fall in the family of Carangidae, Clupeidae, Emmelichthyidae, and Hemiramphidae. -

British Columbia Regional Guide Cat
National Marine Weather Guide British Columbia Regional Guide Cat. No. En56-240/3-2015E-PDF 978-1-100-25953-6 Terms of Usage Information contained in this publication or product may be reproduced, in part or in whole, and by any means, for personal or public non-commercial purposes, without charge or further permission, unless otherwise specified. You are asked to: • Exercise due diligence in ensuring the accuracy of the materials reproduced; • Indicate both the complete title of the materials reproduced, as well as the author organization; and • Indicate that the reproduction is a copy of an official work that is published by the Government of Canada and that the reproduction has not been produced in affiliation with or with the endorsement of the Government of Canada. Commercial reproduction and distribution is prohibited except with written permission from the author. For more information, please contact Environment Canada’s Inquiry Centre at 1-800-668-6767 (in Canada only) or 819-997-2800 or email to [email protected]. Disclaimer: Her Majesty is not responsible for the accuracy or completeness of the information contained in the reproduced material. Her Majesty shall at all times be indemnified and held harmless against any and all claims whatsoever arising out of negligence or other fault in the use of the information contained in this publication or product. Photo credits Cover Left: Chris Gibbons Cover Center: Chris Gibbons Cover Right: Ed Goski Page I: Ed Goski Page II: top left - Chris Gibbons, top right - Matt MacDonald, bottom - André Besson Page VI: Chris Gibbons Page 1: Chris Gibbons Page 5: Lisa West Page 8: Matt MacDonald Page 13: André Besson Page 15: Chris Gibbons Page 42: Lisa West Page 49: Chris Gibbons Page 119: Lisa West Page 138: Matt MacDonald Page 142: Matt MacDonald Acknowledgments Without the works of Owen Lange, this chapter would not have been possible. -

PROXIMATE and GENETIC ANALYSIS of BLACKFIN TUNA (T. Atlanticus)
bioRxiv preprint doi: https://doi.org/10.1101/2020.11.03.366153; this version posted November 4, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. PROXIMATE AND GENETIC ANALYSIS OF BLACKFIN TUNA (T. atlanticus). Yuridia M. Núñez-Mata1, Jesse R. Ríos Rodríguez 1, Adriana L. Perales-Torres 1, Xochitl F. De la Rosa-Reyna2, Jesús A. Vázquez-Rodríguez 3, Nadia A. Fernández-Santos2, Humberto Martínez Montoya 1 * 1 Unidad Académica Multidisciplinaria Reynosa Aztlán – Universidad Autónoma de Tamaulipas. Reynosa, Tamaulipas. 2 Centro de Biotecnología Genómica – Instituto Politécnico Nacional. Reynosa, Tamaulipas. 3 Centro de Investigación en Salud Pública y Nutrición – Universidad Autónoma de Nuevo León. Monterrey, Nuevo León. *Correspondence: [email protected] ORCID: 0000-0003-3228-0054 bioRxiv preprint doi: https://doi.org/10.1101/2020.11.03.366153; this version posted November 4, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. ABSTRACT The tuna meat is a nutritious food that possesses high content of protein, its low content of saturated fatty acids makes it a high demand food in the world. The Thunnus genus is composed of eight species, albacore (T. alalunga), bigeye (T. obesus), long tail tuna (T. tonggol), yellowfin tuna (T. albacares), pacific bluefin tuna (T. orientalis), bluefin tuna (T. maccoyii), Atlantic bluefin tuna ( T. thynnus) and blackfin tuna (T. atlanticus). The blackfin tuna (BFT) (Thunnus atlanticus) represent the smallest species within the Thunnus genus. -

Movements and Diving Behavior of Atlantic Bluefin Tuna Thunnus Thynnus in Relation to Water Column Structure in the Northwestern Atlantic
Vol. 400: 245–265, 2010 MARINE ECOLOGY PROGRESS SERIES Published February 11 doi: 10.3354/meps08394 Mar Ecol Prog Ser Movements and diving behavior of Atlantic bluefin tuna Thunnus thynnus in relation to water column structure in the northwestern Atlantic Gareth L. Lawson1, 2,*, Michael R. Castleton1, Barbara A. Block1 1Tuna Research and Conservation Center, Stanford University, Hopkins Marine Station, 120 Oceanview Boulevard, Pacific Grove, California 93950, USA 2Present address: Biology Department, Woods Hole Oceanographic Institution, Woods Hole, Massachusetts 02543, USA ABSTRACT: We analyzed the movements and diving behavior in relation to water column structure of 35 electronically tagged Atlantic bluefin tuna (176 to 240 cm in length at tagging) during their spring–fall period of occupancy of the Gulf of Maine, Canadian Shelf, and neighboring off-shelf waters from 1999 to 2005. Tagged fish arriving in this study region in March–April initially occupied weakly stratified off-shelf waters along the northern Gulf Stream. As waters over the continental shelf warmed in June, the fish shifted onto the shelf. Sea surface temperatures occupied were relatively constant in both off- and on-shelf waters (April–September monthly medians varying from 16.1 to 19.0°C). Dives made in the stratified waters of the shelf during summer and fall were significantly more frequent (up to 180 dives d–1) and fast (descent rates up to 4.1 m s–1) than in weakly stratified off-shelf waters occupied during spring, defining dives as excursions below tag-derived estimates of the surface isothermal layer depth (ILD). The duration and depth of dives also decreased significantly in associa- tion with changing water column structure, from medians in off-shelf waters during April of 0.45 h and 77.0 m, respectively, to 0.16 h and 24.9 m in August. -

Yellowfin Tuna
Ahi yellown tuna (Thunnus albacares) is one of two Islands. species known in Hawaii simply as Fishing Methods: intermediaries on all islands, or di- ahi. Similar in general appearance rectly to wholesalers and retailers, or it may be shared with family and to bigeye tuna (the other species - known as ahi friends. Most ahi is sold fresh, but men. A large part of the commercial surpluses caught during the peak be recognized by its more torpedo catch (44%) is harvested by longline shaped body, smaller head and eyes. summer season are sometimes dried boats, which may search for tuna and smoked. In Hawaii, shibi is another name up to 800 nautical miles from port and set hooks in deep waters. Yel- Quality to depths below 600 ft. Landings by either bigeye or albacore tuna. Al- lengthen with age. the island of Hawaii, can be sub- stantial (36%) in some years. Troll- Seasonality & How ers contribute most of the remain- does not retain the beautiful natu- They Are Caught der (20%) of the commercial catch ral red color as long as bigeye. The - Availability and Seasonality: - Caught year-round in Hawaii’s wa- ing tournaments held in Hawaii. method, care in handling and other Distribution: abundant during the summer sea- The longline catch and some of the son (May-September). There are handline (ika-shibi) catch of ahi is species. Noticeable changes occur auction. The majority of the hand- Hawaii. ocean surface temperatures and line catch is sold to wholesalers and other oceanographic conditions fa- intermediary buyers on the island of surface during the summer season vor the migration of ahi schools to are susceptible to a quality defect The troll catch may be marketed known as “burnt tuna”. -

MARINE BIRD SURVEYS in QC STRAIT-Final
MARINE BIRD SURVEYS IN QUEEN CHARLOTTE STRAIT AND ADJACENT CHANNELS, AUGUST-SEPTEMBER 2020 December 2020 2 MARINE BIRD SURVEYS IN QUEEN CHARLOTTE STRAIT AND ADJACENT CHANNELS, AUGUST-SEPTEMBER 2020 Anthony Gaston, Mark Maftei, Sonya Pastran, Ken Wright, Graham Sorenson, Iwan Lewylle Cite as: Gaston AJ, M Maftei, S Pastran, K Wright, G Sorenson and I Lewylle. 2020. MARINE BIRD SURVEYS IN QUEEN CHARLOTTE STRAIT AND ADJACENT CHANNELS, AUGUST-SEPTEMBER 2020. Raincoast Education Society, Tofino, BC. 3 1084 Pacific Rim Highway, Tofino, BC (https://raincoasteducation.org/) 4 CONTENTS Introduction 7 Methods Field methods 10 Analysis 13 Results Distribution by zones 16 Total numbers of birds present 17 Timing of arrival/migration Sea ducks 18 Phalaropes 18 Auks 20 Gulls 20 Loons 21 Grebes 22 Petrels 23 Cormorants 23 Comparison with other studies 24 Marine Mammals 26 Discussion Total numbers of birds 28 Timing of arrival/migration 29 Summary 29 Acknowledgements 31 References 32 Appendices 35 5 6 INTRODUCTION Queen Charlotte Strait, a funnel-shaped passage extending ESE from the open waters of Queen Charlotte Sound, separates northern Vancouver Island from the mainland of British Columbia. It is connected, via Johnstone Strait, Discovery Passage and associated channels, with the more or less enclosed waters of the Salish Sea (Figure 1). Anecdotal information from eBird lists suggests that this area supports a high diversity of marine birds in winter and during the periods of northward and southward migration (https://ebird.org/canada/ accessed 7 November 2020; hereafter “eBird”). In addition, this area is important for marine mammals, being used regularly by the Northern Resident Orca stock, and by Humpback and Minke Whales, Dall’s and Harbour Porpoises and Pacific White-sided Dolphins, and supporting several permanent haul-outs of Steller’s Sea Lions (Ford 2014). -

Thunnus Thynnus* the Atlantic Bluefin Tuna
Thunnus Thynnus* The Atlantic Bluefin Tuna An amazing, fishy feat of nature. *Thunnus thynnus. Derived from the Latin “thyno”, “to rush, to dart”. Image courtesy of Chris Gill, Aquila Charters Mevagissey. The return of Atlantic Bluefin Tuna (ABFT) to UK waters in the last few years has generated great interest in this iconic fish from anglers and non-anglers alike. Fishermen will spend thousands and travel across the globe for a chance to hook up to a Bluefin. They are one of the most desirable opponents in the angling world. ABFT are the largest of all the Tuna and can grow to over 700kg, (1,500lbs), and live for up to fifty years. They can attain sppeds of 70kph and dive to depths of over 1000 metres. They undertake great migrations each year, some on a par with those other great ocean travellers, Blue Whales. Their flesh is prized around the world for the finest grades of Sushi, creating a huge global industry that can attach incredible price tags to the best quality fish. Their unique physiology allows them to populate a very wide geographical range. In the Western Atlantic you can find them from the Gulf of Mexico, across the Caribbean, all along the Eastern seaboard of the US, and way up into the cold waters of Canada. In the East, they are found from Morocco, across all of the Mediterranean, and closer to home, Ireland, Wales, Cornwall, Norway, Sweden and Denmark….. Bluefin are almost unique in the geographical and water temperature range of waters they can operate in. -

Psc Draft1 Bc
138°W 136°W 134°W 132°W 130°W 128°W 126°W 124°W 122°W 120°W 118°W N ° 2 6 N ° 2 DR A F T To navigate to PSC Domain 6 1/26/07 maps, click on the legend or on the label on the map. Domain 3: British Columbia R N ° k 0 6 e PSC Region N s ° l Y ukon T 0 e 6 A rritory COBC - Coastal British Columbia Briti sh Columbia FRTH - Fraser R - Thompson R GST - Georgia Strait . JNST - Johnstone Strait R ku NASK - Nass R - Skeena R Ta N QCI - Queen Charlotte Islands ° 8 5 TRAN N TRAN - Transboundary Rivers in Canada ° 8 r 5 ive R WCVI - Western Vancouver Island e in r !. City/Town ik t e v S i Major River R t u k Scale = 1:6,750,000 Is P Miles N ° 0 30 60 120 180 January 2007 6 A B 5 N ° r 6 i 5 C t i s . A h R Alaska l I b C s e F s o a NASK r l N t u a r m I e v S i C tu b R a i rt a N a ° Prince Rupert en!. R 4 ke 5 !. S Terrace iv N e ° r 4 O F 5 !. ras er C H Prince George R e iv c e QCI a r t E e . r R S ate t kw r lac Quesnel A a B !. it D e an R. N C F N COBC h FRTH ° i r 2 lc a 5 o N s ti ° e n !. -

SYNOPSIS on the BIOLOGY of YELLOWFIN TUNA Thunnus (Neothunnus) Albacares (Bonnaterre)1788(PACIFIC OCEAN)
Species Synopsis No. 16 FAO Fisheries Biology Synopsis No, 59 FIb/S59 (Distribution restricted) SAST - Tuna SYNOPSIS ON THE BIOLOGY OF YELLOWFIN TUNA Thunnus (Neothunnus) albacares (Bonnaterre)1788(PACIFIC OCEAN) Exposé synoptique sur la biologie du thon à nageoires jaunes Thunnus (Neothimnus) albacares (Bonnaterre)1788(Océan Pacifique) Sinopsis sobre la. biología dei atítn de aleta amarilla Thunnus (Neothunnus) aibacares (Bonnaterre) 1788 (Ocano Pacífico) Prepared by MILNER B, SCHAEFER, GORDON C,, BROADHEAD and CRAIG J, ORANGE Inter -American Tropical Tuna Commission La Jolla, California, U, S,, A, ISHERIES DIVISION, BIOLOGY BRANCH tOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS R,ome, 1963 538 FIb/S59 Tuna 1:1 i IDENTITY Body plump, wholly covered with scales, which differ in size and form in different parts i. iTaxonomy of the body.Corselet well deveioped but its boundary is not distinct.Tle lateral line has a 1. 1. 1Definition peculiar curve above the pectorals.Teeth rather feeble.Single series of small conical /Fo11owing Berg (1940) modified according to teeth in both jaws.They are sharp and curve Fraser-Brunner (1950)J inward.Villiform teeth on the vomers palatines and pterygoids.Many dentigerous calcareous Phylum VERTEBRATA plates are found on the palate.The denticles on Subphylum Craniata these plates are quite similar to those found on Superclass Gnathostomata the vomer, palatines and pterygoids. Thus the Series Pisces roof of the mouth-cavity is quite rough, contrary Class Teleostomi to the nearly smooth roof in the Katsuwonidae. Subclass Actinopterygii Three lobes of the liver subequal.Intestine Order Perciformes rather long, with three folds.Pyloric tubes Suborder Scombroidei developed only on the posterior convex side of Family Scombridae the duodenum.