Excerpted by Permission from Chapter 10 of the Mistaken Extinction, by Lowell Dingus and Timothy Rowe, New York, W
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Introduction
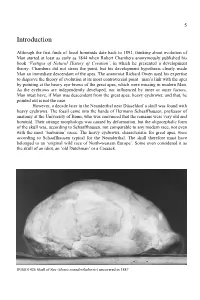
5 Introduction Although the first finds of fossil hominids date back to 1891, thinking about evolution of Man started at least as early as 1844 when Robert Chambers anonymously published his book ‘Vestiges of Natural History of Creation’, in which he presented a development theory. Chambers did not stress the point, but his development hypothesis clearly made Man an immediate descendant of the apes. The anatomist Richard Owen used his expertise to disprove the theory of evolution at its most controversial point –man’s link with the apes by pointing at the heavy eye-brows of the great apes, which were missing in modern Man. As the eyebrows are independently developed, nor influenced by inner or outer factors, Man must have, if Man was descendent from the great apes, heavy eyebrows; and that, he pointed out is not the case. However, a decade later in the Neanderthal near Düsseldorf a skull was found with heavy eyebrows. The fossil came into the hands of Hermann Schaaffhausen, professor of anatomy at the University of Bonn, who was convinced that the remains were very old and hominid. Their strange morphology was caused by deformation, but the oligocephalic form of the skull was, according to Schaaffhausen, not comparable to any modern race, not even with the most ‘barbarian’ races. The heavy eyebrows, characteristic for great apes, were according to Schaaffhausen typical for the Neanderthal. The skull therefore must have belonged to an ‘original wild race of North-western Europe’. Some even considered it as the skull of an idiot, an ‘old Dutchman’ or a Cossack. -

Robert Plot (1641—1696)
( 218 ) SOME EARLY BRITISH ORNITHOLOGISTS AND THEIR WORKS. BY W. H. MULLENS, SI.A., LL.M., M.B.O.U. V.—ROBERT PLOT (1641—1696) AND SOME EARLY COUNTY NATURAL HISTORIES. IN the year 1661, Joshua Childrey (1623—1670), antiquary, schoolmaster, and divine, published in London a small duodecimo work entitled " Britannia Baconia: / or, the Natural / Rarities / of / England, Scotland, & Wales." This book, although of no particular value in itself, being merely a brief and somewhat imperfect compilation, was nevertheless destined to be of some considerable influence on the literature of natural history in this country. For, according to Wood's " Athense Oxonienses" (p. 339), it inspired Robert Plot (1641—1696), the first keeper of the Ashmolean Museum at Oxford, with the idea of writing the " Natural History of Oxfordshire," which appeared in 1677, and which was followed in 1686 by a " Natural History of Staffordshire," the work of the same author ; who is said to have also contemplated similar histories of Middlesex and Kent. These two works of Robert Plot's also proved in their turn to be the forerunners of a numerous series of county natural histories by different writers. The full title of the " Natural History of Oxfordshire" is as follows :— " The / Natural History / of / Oxford-shire, / being an Essay toward the Natural History / of / England. / By R.P., LL.D. / [quotation from Arat. in Phsenom.] / [engraving] Printed at the Theater in Oxford, and are to be had there: / And in London at Mr. S. Millers, at the Star near the / West-end of SOME EARLY BRITISH ORNITHOLOGISTS. -

The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886
The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886 Sascha Nolden, Simon Nathan & Esme Mildenhall Geoscience Society of New Zealand miscellaneous publication 133H November 2013 Published by the Geoscience Society of New Zealand Inc, 2013 Information on the Society and its publications is given at www.gsnz.org.nz © Copyright Simon Nathan & Sascha Nolden, 2013 Geoscience Society of New Zealand miscellaneous publication 133H ISBN 978-1-877480-29-4 ISSN 2230-4495 (Online) ISSN 2230-4487 (Print) We gratefully acknowledge financial assistance from the Brian Mason Scientific and Technical Trust which has provided financial support for this project. This document is available as a PDF file that can be downloaded from the Geoscience Society website at: http://www.gsnz.org.nz/information/misc-series-i-49.html Bibliographic Reference Nolden, S.; Nathan, S.; Mildenhall, E. 2013: The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886. Geoscience Society of New Zealand miscellaneous publication 133H. 219 pages. The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886 CONTENTS Introduction 3 The Sumner Cave controversy Sources of the Haast-Hooker correspondence Transcription and presentation of the letters Acknowledgements References Calendar of Letters 8 Transcriptions of the Haast-Hooker letters 12 Appendix 1: Undated letter (fragment), ca 1867 208 Appendix 2: Obituary for Sir Julius von Haast 209 Appendix 3: Biographical register of names mentioned in the correspondence 213 Figures Figure 1: Photographs -

Chapter 10, the Mistaken Extinction, by Lowell Dingus and Timothy Rowe, New York, W
Chapter 10, The Mistaken Extinction, by Lowell Dingus and Timothy Rowe, New York, W. H. Freeman, 1998. CHAPTER 10 Dinosaurs Challenge Evolution Enter Sir Richard Owen More than 150 years ago, the great British naturalist Richard Owen (fig. 10.01) ignited the controversy that Deinonychus would eventually inflame. The word "dinosaur" was first uttered by Owen in a lecture delivered at Plymouth, England in July of 1841. He had coined the name in a report on giant fossil reptiles that were discovered in England earlier in the century. The root, Deinos, is usually translated as "terrible" but in his report, published in 1842, Owen chose the words "fearfully great"1. To Owen, dinosaurs were the fearfully great saurian reptiles, known only from fossil skeletons of huge extinct animals, unlike anything alive today. Fig. 10.01 Richard Owen as, A) a young man at about the time he named Dinosauria, B) in middle age, near the time he described Archaeopteryx, and C) in old age. Dinosaur bones were discovered long before Owen first spoke their name, but no one understood what they represented. The first scientific report on a dinosaur bone belonging was printed in 1677 by Rev. Robert Plot in his work, The Natural History of Oxfordshire. This broken end of a thigh bone, came to Plot's attention during his research. It was nearly 60 cm in circumference--greater than the same bone in an elephant (fig.10.02). We now suspect that it belonged to Megalosaurus bucklandii, a carnivorous dinosaur now known from Oxfordshire. But Plot concluded that it "must have been a real Bone, now petrified" and that it resembled "exactly the figure of the 1 Chapter 10, The Mistaken Extinction, by Lowell Dingus and Timothy Rowe, New York, W. -

Natural History of Staffordshire Printed in Oxford in 1686, the Subject of This Article
THE NATURAL HISTORY OF STAFFORD-SHIRE. By Robert Plot, LLD. Keeper of the Ashmolean Musæum and Professor of Chymistry in the University of Oxford. Ye shall Describe the Land, and bring the Description hither to Me. Joshua 8. v. 6. Oxford printed at the Theatre, Anno M. DC. LXXXVI. (1686) by Yasha Beresiner Introduction This volume, now in the Masonic Library of the Regular Grand Lodge of Italy, contains the earliest recorded account of accepted masonry and is considered the most implicit report on the fraternity available for the period at the end of the 17th century. It is printed in paragraphs 85 to 88 inclusive, on pages 316 to 318 of the tome. This text is referred to as the Plot Abstract. Its importance lies a) with regard to its content i.e. the summary of the legendary history, the description of contemporary freemasonry, and the criticisms of the fraternity and b) as to the sources from which Plot may have derived his information, most importantly, what he refers to as the ‘large parchment volum they have amongst them . .’ The purpose of this article is not to analyse the text and its content but rather to identify and clarify the reasons behind the importance of this Volume in the context of Masonic bibliography and history. Robert Plot Robert Plot (1640-96) was born in Kent and received a BA degree from Oxford University in 1661, an MA in 1664 and a Law degrees in 1671. In 1677 he was elected a Fellow of the Royal Society and became Secretary in 1682. -

New Perspectives on Pterosaur Palaeobiology
Downloaded from http://sp.lyellcollection.org/ by guest on September 24, 2021 New perspectives on pterosaur palaeobiology DAVID W. E. HONE1*, MARK P. WITTON2 & DAVID M. MARTILL2 1Queen Mary University of London, Mile End Road, London E14NS, UK 2School of Earth and Environmental Sciences, University of Portsmouth, Burnaby Road, Portsmouth PO1 3QL, UK *Correspondence: [email protected] Abstract: Pterosaurs were the first vertebrates to evolve powered flight and occupied the skies of the Mesozoic for 160 million years. They occurred on every continent, evolved their incredible pro- portions and anatomy into well over 100 species, and included the largest flying animals of all time among their ranks. Pterosaurs are undergoing a long-running scientific renaissance that has seen elevated interest from a new generation of palaeontologists, contributions from scientists working all over the world and major advances in our understanding of their palaeobiology. They have espe- cially benefited from the application of new investigative techniques applied to historical speci- mens and the discovery of new material, including detailed insights into their fragile skeletons and their soft tissue anatomy. Many aspects of pterosaur science remain controversial, mainly due to the investigative challenges presented by their fragmentary, fragile fossils and notoriously patchy fossil record. With perseverance, these controversies are being resolved and our understand- ing of flying reptiles is increasing. This volume brings together a diverse set of papers on numerous aspects of the biology of these fascinating reptiles, including discussions of pterosaur ecology, flight, ontogeny, bony and soft tissue anatomy, distribution and evolution, as well as revisions of their taxonomy and relationships. -

Designing the Dinosaur: Richard Owen's Response to Robert Edmond Grant Author(S): Adrian J
Designing the Dinosaur: Richard Owen's Response to Robert Edmond Grant Author(s): Adrian J. Desmond Source: Isis, Vol. 70, No. 2 (Jun., 1979), pp. 224-234 Published by: The University of Chicago Press on behalf of The History of Science Society Stable URL: http://www.jstor.org/stable/230789 . Accessed: 16/10/2013 13:00 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. The University of Chicago Press and The History of Science Society are collaborating with JSTOR to digitize, preserve and extend access to Isis. http://www.jstor.org This content downloaded from 150.135.115.18 on Wed, 16 Oct 2013 13:00:27 PM All use subject to JSTOR Terms and Conditions Designing the Dinosaur: Richard Owen's Response to Robert Edmond Grant By Adrian J. Desmond* I N THEIR PAPER on "The Earliest Discoveries of Dinosaurs" Justin Delair and William Sarjeant permit Richard Owen to step in at the last moment and cap two decades of frenzied fossil collecting with the word "dinosaur."' This approach, I believe, denies Owen's real achievement while leaving a less than fair impression of the creative aspect of science. -

Marsupials As Ancestors Or Sister Taxa?
Archives of natural history 39.2 (2012): 217–233 Edinburgh University Press DOI: 10.3366/anh.2012.0091 # The Society for the History of Natural History www.eupjournals.com/anh Darwin’s two competing phylogenetic trees: marsupials as ancestors or sister taxa? J. DAVID ARCHIBALD Department of Biology, San Diego State University, San Diego, CA 92182–4614, USA (e-mail: [email protected]). ABSTRACT: Studies of the origin and diversification of major groups of plants and animals are contentious topics in current evolutionary biology. This includes the study of the timing and relationships of the two major clades of extant mammals – marsupials and placentals. Molecular studies concerned with marsupial and placental origin and diversification can be at odds with the fossil record. Such studies are, however, not a recent phenomenon. Over 150 years ago Charles Darwin weighed two alternative views on the origin of marsupials and placentals. Less than a year after the publication of On the origin of species, Darwin outlined these in a letter to Charles Lyell dated 23 September 1860. The letter concluded with two competing phylogenetic diagrams. One showed marsupials as ancestral to both living marsupials and placentals, whereas the other showed a non-marsupial, non-placental as being ancestral to both living marsupials and placentals. These two diagrams are published here for the first time. These are the only such competing phylogenetic diagrams that Darwin is known to have produced. In addition to examining the question of mammalian origins in this letter and in other manuscript notes discussed here, Darwin confronted the broader issue as to whether major groups of animals had a single origin (monophyly) or were the result of “continuous creation” as advocated for some groups by Richard Owen. -

Phylogeny and Avian Evolution Phylogeny and Evolution of the Aves
Phylogeny and Avian Evolution Phylogeny and Evolution of the Aves I. Background Scientists have speculated about evolution of birds ever since Darwin. Difficult to find relatives using only modern animals After publi cati on of “O rigi i in of S peci es” (~1860) some used birds as a counter-argument since th ere were no k nown t ransiti onal f orms at the time! • turtles have modified necks and toothless beaks • bats fly and are warm blooded With fossil discovery other potential relationships! • Birds as distinct order of reptiles Many non-reptilian characteristics (e.g. endothermy, feathers) but really reptilian in structure! If birds only known from fossil record then simply be a distinct order of reptiles. II. Reptile Evolutionary History A. “Stem reptiles” - Cotylosauria Must begin in the late Paleozoic ClCotylosauri a – “il”“stem reptiles” Radiation of reptiles from Cotylosauria can be organized on the basis of temporal fenestrae (openings in back of skull for muscle attachment). Subsequent reptilian lineages developed more powerful jaws. B. Anapsid Cotylosauria and Chelonia have anapsid pattern C. Syypnapsid – single fenestra Includes order Therapsida which gave rise to mammalia D. Diapsida – both supppratemporal and infratemporal fenestrae PttPattern foun did in exti titnct arch osaurs, survi iiving archosaurs and also in primitive lepidosaur – ShSpheno don. All remaining living reptiles and the lineage leading to Aves are classified as Diapsida Handout Mammalia Extinct Groups Cynodontia Therapsida Pelycosaurs Lepidosauromorpha Ichthyosauria Protorothyrididae Synapsida Anapsida Archosauromorpha Euryapsida Mesosaurs Amphibia Sauria Diapsida Eureptilia Sauropsida Amniota Tetrapoda III. Relationshippp to Reptiles Most groups present during Mesozoic considere d ancestors to bird s. -

BIOL 1406 Darwin's Dangerous Idea
BIOL 1406 Darwin’s Dangerous Idea - Video Exam I Essay Question: (Matching Format) Describe the history of the scientific theory, biological evolution by means of natural selection: and focus on the life of Charles Darwin as portrayed in the PBS production, Darwin’s Dangerous Idea. Be sure to describe the roles of the following: "Raz", Robert FitzRoy, Emma Darwin, Annie Darwin, Richard Owen, Charles Lyell, Thomas Malthus, Samuel Wilberforce, and Thomas Huxley.) 1.Describe Captain Fitzroy’s perspective when it comes to “free-thinking” 2. What does Fitzroy allow Darwin to borrow? 3. Who was “Raz”? 4. Who was Richard Owen? 5. What was Owen’s view on “free-thinking” with regard to human ancestory? 6. What was Owen so afraid of? 7. Who was Emma (Wedgewood) Darwin? How did she influence Charles Darwin with regard to his scientific inquiry ? 8. What type of disease do we now speculate that Darwin may have suffered from? How did he get the disease? 9. Who was Annie Darwin? 10. When Annie left, what affect did this have on Darwin? 11. Who was Charles Lyell? What role did he play in influencing Darwin? 12. Who was Thomas Malthus? What did he do to influence Darwin? 13. What did Richard Owen do that was scientifically unethical? Why did he do this? 14. Who was Samuel Wilberforce? 15. Who was Thomas Henry Huxley? What did he do to influence Darwin? 16. Who was Alfred Russel Wallace? What did he do to influence Darwin? 17. What motivated Darwin to study so many different organisms; i.e. -

Styles of Reasoning in Early to Mid-Victorian Life Research: Analysis:Synthesis and Palaetiology
Journal of the History of Biology (2006) Ó Springer 2006 DOI 10.1007/s10739-006-0006-4 Styles of Reasoning in Early to Mid-Victorian Life Research: Analysis:Synthesis and Palaetiology JAMES ELWICK Science and Technology Studies Faculties of Arts and Science and Engineering York University 4700 Keele St. M3J 1P3 Toronto, ON Canada E-mail: [email protected] Abstract. To better understand the work of pre-Darwinian British life researchers in their own right, this paper discusses two different styles of reasoning. On the one hand there was analysis:synthesis, where an organism was disintegrated into its constituent parts and then reintegrated into a whole; on the other hand there was palaetiology, the historicist depiction of the progressive specialization of an organism. This paper shows how each style allowed for development, but showed it as moving in opposite directions. In analysis:synthesis, development proceeded centripetally, through the fusion of parts. Meanwhile in palaetiology, development moved centrifugally, through the ramifying specialization of an initially simple substance. I first examine a com- munity of analytically oriented British life researchers, exemplified by Richard Owen, and certain technical questions they considered important. These involved the neu- rosciences, embryology, and reproduction and regeneration. The paper then looks at a new generation of British palaetiologists, exemplified by W.B. Carpenter and T.H. Huxley, who succeeded at portraying analysts’ questions as irrelevant. The link between styles of reasoning and physical sites is also explored. Analysts favored museums, which facilitated the examination and display of unchanging marine organisms while providing a power base for analysts. I suggest that palaetiologists were helped by vivaria, which included marine aquaria and Wardian cases. -

How to Find a Dinosaur, and the Role of Synonymy in Biodiversity Studies
Paleobiology, 34(4), 2008, pp. 516–533 How to find a dinosaur, and the role of synonymy in biodiversity studies Michael J. Benton Abstract.—Taxon discovery underlies many studies in evolutionary biology, including biodiversity and conservation biology. Synonymy has been recognized as an issue, and as many as 30–60% of named species later turn out to be invalid as a result of synonymy or other errors in taxonomic practice. This error level cannot be ignored, because users of taxon lists do not know whether their data sets are clean or riddled with erroneous taxa. A year-by-year study of a large clade, Dinosauria, comprising over 1000 taxa, reveals how systematists have worked. The group has been subject to heavy review and revision over the decades, and the error rate is about 40% at generic level and 50% at species level. The naming of new species and genera of dinosaurs is proportional to the number of people at work in the field. But the number of valid new dinosaurian taxa depends mainly on the discovery of new territory, particularly new sedimentary basins, as well as the number of paleontologists. Error rates are highest (Ͼ50%) for dinosaurs from Europe; less well studied con- tinents show lower totals of taxa, exponential discovery curves, and lower synonymy rates. The most prolific author of new dinosaur names was Othniel Marsh, who named 80 species, closely followed by Friedrich von Huene (71) and Edward Cope (64), but the ‘‘success rate’’ (proportion of dinosaurs named that are still regarded as valid) was low (0.14–0.29) for these earlier authors, and it appears to improve through time, partly a reflection of reduction in revision time, but mainly because modern workers base their new taxa on more complete specimens.