Marsupials As Ancestors Or Sister Taxa?
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Musky Rat-Kangaroos, Hypsiprymnodon Moschatus: Cursorial Frugivores in Australia's Wet-Tropical Rain Forests
ResearchOnline@JCU This file is part of the following reference: Dennis, Andrew James (1997) Musky Rat-kangaroos, hypsiprymnodon moschatus: cursorial frugivores in Australia's wet-tropical rain forests. PhD thesis, James Cook University. Access to this file is available from: http://eprints.jcu.edu.au/17401/ If you believe that this work constitutes a copyright infringement, please contact [email protected] and quote http://eprints.jcu.edu.au/17401/ Chapter 11 MUSKY RAT-KANGAROOS: CURSORIAL FRUGIVORES How do Musky Rat-kangaroos Relate !Q..their Environment ? Musky Rat-kangaroos can be classified as frugivo res because fr ui ts and seeds accounted fo r the bul k of their diet th roughout the year (Chapter 3). In addi tion, they scatterhoarded many fruits and seeds, to the benefit of at least some species of plants (Chapters 8 & 9). They consumed most of the available frui ts which had a fleshy pericarp or ani but also included the seeds of some species that did not. They ate fruits from over half the species producing fruits on my study site, many of those they did not eat were wind dispersed, housed in hard, indehiscent pods or had furry. dehiscent pods. In addition, some fleshy drupes were not consumed. Like many other frugivores, Musky Rat-kangaroos supplemented their diet from other sources, particularly when the seasonal availabil ity of fruits is at its min imum (see Terborgh 1983). During the late wet, co ld an d early dry seasons, when frui t abundance was mini mal, their search effort was random with respect to fr ui t fall s (Figure 3.8; Table 3.2). -
Introduction
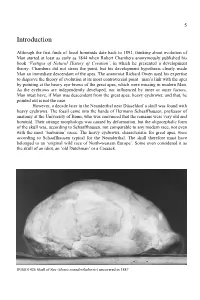
5 Introduction Although the first finds of fossil hominids date back to 1891, thinking about evolution of Man started at least as early as 1844 when Robert Chambers anonymously published his book ‘Vestiges of Natural History of Creation’, in which he presented a development theory. Chambers did not stress the point, but his development hypothesis clearly made Man an immediate descendant of the apes. The anatomist Richard Owen used his expertise to disprove the theory of evolution at its most controversial point –man’s link with the apes by pointing at the heavy eye-brows of the great apes, which were missing in modern Man. As the eyebrows are independently developed, nor influenced by inner or outer factors, Man must have, if Man was descendent from the great apes, heavy eyebrows; and that, he pointed out is not the case. However, a decade later in the Neanderthal near Düsseldorf a skull was found with heavy eyebrows. The fossil came into the hands of Hermann Schaaffhausen, professor of anatomy at the University of Bonn, who was convinced that the remains were very old and hominid. Their strange morphology was caused by deformation, but the oligocephalic form of the skull was, according to Schaaffhausen, not comparable to any modern race, not even with the most ‘barbarian’ races. The heavy eyebrows, characteristic for great apes, were according to Schaaffhausen typical for the Neanderthal. The skull therefore must have belonged to an ‘original wild race of North-western Europe’. Some even considered it as the skull of an idiot, an ‘old Dutchman’ or a Cossack. -

The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886
The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886 Sascha Nolden, Simon Nathan & Esme Mildenhall Geoscience Society of New Zealand miscellaneous publication 133H November 2013 Published by the Geoscience Society of New Zealand Inc, 2013 Information on the Society and its publications is given at www.gsnz.org.nz © Copyright Simon Nathan & Sascha Nolden, 2013 Geoscience Society of New Zealand miscellaneous publication 133H ISBN 978-1-877480-29-4 ISSN 2230-4495 (Online) ISSN 2230-4487 (Print) We gratefully acknowledge financial assistance from the Brian Mason Scientific and Technical Trust which has provided financial support for this project. This document is available as a PDF file that can be downloaded from the Geoscience Society website at: http://www.gsnz.org.nz/information/misc-series-i-49.html Bibliographic Reference Nolden, S.; Nathan, S.; Mildenhall, E. 2013: The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886. Geoscience Society of New Zealand miscellaneous publication 133H. 219 pages. The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886 CONTENTS Introduction 3 The Sumner Cave controversy Sources of the Haast-Hooker correspondence Transcription and presentation of the letters Acknowledgements References Calendar of Letters 8 Transcriptions of the Haast-Hooker letters 12 Appendix 1: Undated letter (fragment), ca 1867 208 Appendix 2: Obituary for Sir Julius von Haast 209 Appendix 3: Biographical register of names mentioned in the correspondence 213 Figures Figure 1: Photographs -

Constraints on the Timescale of Animal Evolutionary History
Palaeontologia Electronica palaeo-electronica.org Constraints on the timescale of animal evolutionary history Michael J. Benton, Philip C.J. Donoghue, Robert J. Asher, Matt Friedman, Thomas J. Near, and Jakob Vinther ABSTRACT Dating the tree of life is a core endeavor in evolutionary biology. Rates of evolution are fundamental to nearly every evolutionary model and process. Rates need dates. There is much debate on the most appropriate and reasonable ways in which to date the tree of life, and recent work has highlighted some confusions and complexities that can be avoided. Whether phylogenetic trees are dated after they have been estab- lished, or as part of the process of tree finding, practitioners need to know which cali- brations to use. We emphasize the importance of identifying crown (not stem) fossils, levels of confidence in their attribution to the crown, current chronostratigraphic preci- sion, the primacy of the host geological formation and asymmetric confidence intervals. Here we present calibrations for 88 key nodes across the phylogeny of animals, rang- ing from the root of Metazoa to the last common ancestor of Homo sapiens. Close attention to detail is constantly required: for example, the classic bird-mammal date (base of crown Amniota) has often been given as 310-315 Ma; the 2014 international time scale indicates a minimum age of 318 Ma. Michael J. Benton. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Philip C.J. Donoghue. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Robert J. -

Ba3444 MAMMAL BOOKLET FINAL.Indd
Intot Obliv i The disappearing native mammals of northern Australia Compiled by James Fitzsimons Sarah Legge Barry Traill John Woinarski Into Oblivion? The disappearing native mammals of northern Australia 1 SUMMARY Since European settlement, the deepest loss of Australian biodiversity has been the spate of extinctions of endemic mammals. Historically, these losses occurred mostly in inland and in temperate parts of the country, and largely between 1890 and 1950. A new wave of extinctions is now threatening Australian mammals, this time in northern Australia. Many mammal species are in sharp decline across the north, even in extensive natural areas managed primarily for conservation. The main evidence of this decline comes consistently from two contrasting sources: robust scientifi c monitoring programs and more broad-scale Indigenous knowledge. The main drivers of the mammal decline in northern Australia include inappropriate fi re regimes (too much fi re) and predation by feral cats. Cane Toads are also implicated, particularly to the recent catastrophic decline of the Northern Quoll. Furthermore, some impacts are due to vegetation changes associated with the pastoral industry. Disease could also be a factor, but to date there is little evidence for or against it. Based on current trends, many native mammals will become extinct in northern Australia in the next 10-20 years, and even the largest and most iconic national parks in northern Australia will lose native mammal species. This problem needs to be solved. The fi rst step towards a solution is to recognise the problem, and this publication seeks to alert the Australian community and decision makers to this urgent issue. -

71St Annual Meeting Society of Vertebrate Paleontology Paris Las Vegas Las Vegas, Nevada, USA November 2 – 5, 2011 SESSION CONCURRENT SESSION CONCURRENT
ISSN 1937-2809 online Journal of Supplement to the November 2011 Vertebrate Paleontology Vertebrate Society of Vertebrate Paleontology Society of Vertebrate 71st Annual Meeting Paleontology Society of Vertebrate Las Vegas Paris Nevada, USA Las Vegas, November 2 – 5, 2011 Program and Abstracts Society of Vertebrate Paleontology 71st Annual Meeting Program and Abstracts COMMITTEE MEETING ROOM POSTER SESSION/ CONCURRENT CONCURRENT SESSION EXHIBITS SESSION COMMITTEE MEETING ROOMS AUCTION EVENT REGISTRATION, CONCURRENT MERCHANDISE SESSION LOUNGE, EDUCATION & OUTREACH SPEAKER READY COMMITTEE MEETING POSTER SESSION ROOM ROOM SOCIETY OF VERTEBRATE PALEONTOLOGY ABSTRACTS OF PAPERS SEVENTY-FIRST ANNUAL MEETING PARIS LAS VEGAS HOTEL LAS VEGAS, NV, USA NOVEMBER 2–5, 2011 HOST COMMITTEE Stephen Rowland, Co-Chair; Aubrey Bonde, Co-Chair; Joshua Bonde; David Elliott; Lee Hall; Jerry Harris; Andrew Milner; Eric Roberts EXECUTIVE COMMITTEE Philip Currie, President; Blaire Van Valkenburgh, Past President; Catherine Forster, Vice President; Christopher Bell, Secretary; Ted Vlamis, Treasurer; Julia Clarke, Member at Large; Kristina Curry Rogers, Member at Large; Lars Werdelin, Member at Large SYMPOSIUM CONVENORS Roger B.J. Benson, Richard J. Butler, Nadia B. Fröbisch, Hans C.E. Larsson, Mark A. Loewen, Philip D. Mannion, Jim I. Mead, Eric M. Roberts, Scott D. Sampson, Eric D. Scott, Kathleen Springer PROGRAM COMMITTEE Jonathan Bloch, Co-Chair; Anjali Goswami, Co-Chair; Jason Anderson; Paul Barrett; Brian Beatty; Kerin Claeson; Kristina Curry Rogers; Ted Daeschler; David Evans; David Fox; Nadia B. Fröbisch; Christian Kammerer; Johannes Müller; Emily Rayfield; William Sanders; Bruce Shockey; Mary Silcox; Michelle Stocker; Rebecca Terry November 2011—PROGRAM AND ABSTRACTS 1 Members and Friends of the Society of Vertebrate Paleontology, The Host Committee cordially welcomes you to the 71st Annual Meeting of the Society of Vertebrate Paleontology in Las Vegas. -

Chapter 10, the Mistaken Extinction, by Lowell Dingus and Timothy Rowe, New York, W
Chapter 10, The Mistaken Extinction, by Lowell Dingus and Timothy Rowe, New York, W. H. Freeman, 1998. CHAPTER 10 Dinosaurs Challenge Evolution Enter Sir Richard Owen More than 150 years ago, the great British naturalist Richard Owen (fig. 10.01) ignited the controversy that Deinonychus would eventually inflame. The word "dinosaur" was first uttered by Owen in a lecture delivered at Plymouth, England in July of 1841. He had coined the name in a report on giant fossil reptiles that were discovered in England earlier in the century. The root, Deinos, is usually translated as "terrible" but in his report, published in 1842, Owen chose the words "fearfully great"1. To Owen, dinosaurs were the fearfully great saurian reptiles, known only from fossil skeletons of huge extinct animals, unlike anything alive today. Fig. 10.01 Richard Owen as, A) a young man at about the time he named Dinosauria, B) in middle age, near the time he described Archaeopteryx, and C) in old age. Dinosaur bones were discovered long before Owen first spoke their name, but no one understood what they represented. The first scientific report on a dinosaur bone belonging was printed in 1677 by Rev. Robert Plot in his work, The Natural History of Oxfordshire. This broken end of a thigh bone, came to Plot's attention during his research. It was nearly 60 cm in circumference--greater than the same bone in an elephant (fig.10.02). We now suspect that it belonged to Megalosaurus bucklandii, a carnivorous dinosaur now known from Oxfordshire. But Plot concluded that it "must have been a real Bone, now petrified" and that it resembled "exactly the figure of the 1 Chapter 10, The Mistaken Extinction, by Lowell Dingus and Timothy Rowe, New York, W. -

Designing the Dinosaur: Richard Owen's Response to Robert Edmond Grant Author(S): Adrian J
Designing the Dinosaur: Richard Owen's Response to Robert Edmond Grant Author(s): Adrian J. Desmond Source: Isis, Vol. 70, No. 2 (Jun., 1979), pp. 224-234 Published by: The University of Chicago Press on behalf of The History of Science Society Stable URL: http://www.jstor.org/stable/230789 . Accessed: 16/10/2013 13:00 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. The University of Chicago Press and The History of Science Society are collaborating with JSTOR to digitize, preserve and extend access to Isis. http://www.jstor.org This content downloaded from 150.135.115.18 on Wed, 16 Oct 2013 13:00:27 PM All use subject to JSTOR Terms and Conditions Designing the Dinosaur: Richard Owen's Response to Robert Edmond Grant By Adrian J. Desmond* I N THEIR PAPER on "The Earliest Discoveries of Dinosaurs" Justin Delair and William Sarjeant permit Richard Owen to step in at the last moment and cap two decades of frenzied fossil collecting with the word "dinosaur."' This approach, I believe, denies Owen's real achievement while leaving a less than fair impression of the creative aspect of science. -

Wanburoo Hilarus Gen. Et Sp. Nov., a Lophodont Bulungamayine Kangaroo
Records of the Western Australian Museum Supplement No. 57: 239-253 (1999). Wanburoo hilarus gen. et sp. nov., a lophodont bulungamayine kangaroo (Marsupialia: Macropodoidea: Bulungamayinae) from the Miocene deposits of Riversleigh, northwestern Queensland Bernard N. Cooke School of Life Sciences, Queensland University of Technology, GPO Box 2434, Brisbane, Qld 4001; email: [email protected] and, School of Biological Science, University of New South Wales, Sydney, NSW 2052 Abstract - Wanburoo hi/ants gen. et sp. novo is described on the basis of specimens recovered from a number of Riversleigh sites ranging in estimated age from early Middle Miocene to early Late Miocene. The new species is characterized by low-crowned, lophodont molars in which hypolophid morphology clearly indicates bulungamayine affinity. The relatively close temporal proximity and lophodont molar morphology of the new species invites comparison with the Late Miocene macropodids, Dorcopsoides fossilis and Hadronomas puckridgi. While there is a number of phenetic similarities between the species, many of these appear to be symplesiomorphic. However, a number of apomorphies are suggested as indicating a phylogenetic relationship between the new species and early, non-balbarine macropodids represented by D. fossilis and H. puckridgi. Of these two taxa, the relationship appears stronger with Hadronomas than with Dorcopsoides. Balbarine-like characters present in Dorcopsoides are likely plesiomorphic or the result of convergence. While bulungamayines are herein regarded as more likely than balbarines to be ancestral to macropodids, the possibility of a diphyletic origin for macropodids cannot be dismissed. The suggested relationship between bulungamayines and macropodids casts doubt on the inclusion of Bulungamayinae within Potoroidae. -

BIOL 1406 Darwin's Dangerous Idea
BIOL 1406 Darwin’s Dangerous Idea - Video Exam I Essay Question: (Matching Format) Describe the history of the scientific theory, biological evolution by means of natural selection: and focus on the life of Charles Darwin as portrayed in the PBS production, Darwin’s Dangerous Idea. Be sure to describe the roles of the following: "Raz", Robert FitzRoy, Emma Darwin, Annie Darwin, Richard Owen, Charles Lyell, Thomas Malthus, Samuel Wilberforce, and Thomas Huxley.) 1.Describe Captain Fitzroy’s perspective when it comes to “free-thinking” 2. What does Fitzroy allow Darwin to borrow? 3. Who was “Raz”? 4. Who was Richard Owen? 5. What was Owen’s view on “free-thinking” with regard to human ancestory? 6. What was Owen so afraid of? 7. Who was Emma (Wedgewood) Darwin? How did she influence Charles Darwin with regard to his scientific inquiry ? 8. What type of disease do we now speculate that Darwin may have suffered from? How did he get the disease? 9. Who was Annie Darwin? 10. When Annie left, what affect did this have on Darwin? 11. Who was Charles Lyell? What role did he play in influencing Darwin? 12. Who was Thomas Malthus? What did he do to influence Darwin? 13. What did Richard Owen do that was scientifically unethical? Why did he do this? 14. Who was Samuel Wilberforce? 15. Who was Thomas Henry Huxley? What did he do to influence Darwin? 16. Who was Alfred Russel Wallace? What did he do to influence Darwin? 17. What motivated Darwin to study so many different organisms; i.e. -

Styles of Reasoning in Early to Mid-Victorian Life Research: Analysis:Synthesis and Palaetiology
Journal of the History of Biology (2006) Ó Springer 2006 DOI 10.1007/s10739-006-0006-4 Styles of Reasoning in Early to Mid-Victorian Life Research: Analysis:Synthesis and Palaetiology JAMES ELWICK Science and Technology Studies Faculties of Arts and Science and Engineering York University 4700 Keele St. M3J 1P3 Toronto, ON Canada E-mail: [email protected] Abstract. To better understand the work of pre-Darwinian British life researchers in their own right, this paper discusses two different styles of reasoning. On the one hand there was analysis:synthesis, where an organism was disintegrated into its constituent parts and then reintegrated into a whole; on the other hand there was palaetiology, the historicist depiction of the progressive specialization of an organism. This paper shows how each style allowed for development, but showed it as moving in opposite directions. In analysis:synthesis, development proceeded centripetally, through the fusion of parts. Meanwhile in palaetiology, development moved centrifugally, through the ramifying specialization of an initially simple substance. I first examine a com- munity of analytically oriented British life researchers, exemplified by Richard Owen, and certain technical questions they considered important. These involved the neu- rosciences, embryology, and reproduction and regeneration. The paper then looks at a new generation of British palaetiologists, exemplified by W.B. Carpenter and T.H. Huxley, who succeeded at portraying analysts’ questions as irrelevant. The link between styles of reasoning and physical sites is also explored. Analysts favored museums, which facilitated the examination and display of unchanging marine organisms while providing a power base for analysts. I suggest that palaetiologists were helped by vivaria, which included marine aquaria and Wardian cases. -

Miranda, 5 | 2011 Jim Endersby, Imperial Nature: Joseph Hooker and the Practices of Victorian S
Miranda Revue pluridisciplinaire du monde anglophone / Multidisciplinary peer-reviewed journal on the English- speaking world 5 | 2011 South and Race / Staging Mobility in the United States Jim Endersby, Imperial Nature: Joseph Hooker and the Practices of Victorian Science Laurence Talairach-Vielmas Édition électronique URL : http://journals.openedition.org/miranda/2550 ISSN : 2108-6559 Éditeur Université Toulouse - Jean Jaurès Référence électronique Laurence Talairach-Vielmas, « Jim Endersby, Imperial Nature: Joseph Hooker and the Practices of Victorian Science », Miranda [En ligne], 5 | 2011, mis en ligne le 29 novembre 2011, consulté le 25 octobre 2018. URL : http://journals.openedition.org/miranda/2550 Ce document a été généré automatiquement le 25 octobre 2018. Miranda is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License. Jim Endersby, Imperial Nature: Joseph Hooker and the Practices of Victorian S... 1 Jim Endersby, Imperial Nature: Joseph Hooker and the Practices of Victorian Science Laurence Talairach-Vielmas RÉFÉRENCE Jim Endersby, Imperial Nature : Joseph Hooker and the Practices of Victorian Science (Chicago and London : The University of Chicago Press, [2008] 2010), 429 p, ISBN 978–0–226–20791– 9 1 L’ouvrage de Jim Endersby, Imperial Nature : Joseph Hooker and the Practices of Victorian Science, ne se veut pas une biographie du naturaliste Joseph Dalton Hooker (1817–1911). Bien au contraire. Hooker fut un scientifique véritablement victorien, dont la carrière retrace l’évolution du statut de scientifique, les enjeux et tensions au cœur de la profession et les liens entre scientifiques et naturalistes amateurs. L’étude de Endersby se concentre sur les années charnières de la carrière de Hooker, avant son accession à la tête de Kew Gardens, à un moment où le statut du scientifique est en pleine évolution.