Oceanography
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Fishes of the Eastern Ross Sea, Antarctica
Polar Biol (2004) 27: 637–650 DOI 10.1007/s00300-004-0632-2 REVIEW Joseph Donnelly Æ Joseph J. Torres Tracey T. Sutton Æ Christina Simoniello Fishes of the eastern Ross Sea, Antarctica Received: 26 November 2003 / Revised: 16 April 2004 / Accepted: 20 April 2004 / Published online: 16 June 2004 Ó Springer-Verlag 2004 Abstract Antarctic fishes were sampled with 41 midwater in Antarctica is dominated by a few fish families and 6 benthic trawls during the 1999–2000 austral (Bathylagidae, Gonostomatidae, Myctophidae and summer in the eastern Ross Sea. The oceanic pelagic Paralepididae) with faunal diversity decreasing south assemblage (0–1,000 m) contained Electrona antarctica, from the Antarctic Polar Front to the continent (Ever- Gymnoscopelus opisthopterus, Bathylagus antarcticus, son 1984; Kock 1992; Kellermann 1996). South of the Cyclothone kobayashii and Notolepis coatsi. These were Polar Front, the majority of meso- and bathypelagic replaced over the shelf by notothenioids, primarily Ple- fishes have circum-Antarctic distributions (McGinnis uragramma antarcticum. Pelagic biomass was low and 1982; Gon and Heemstra 1990). Taken collectively, the concentrated below 500 m. The demersal assemblage fishes are significant contributors to the pelagic biomass was characteristic of East Antarctica and included seven and are important trophic elements, both as predators species each of Artedidraconidae, Bathydraconidae and and prey (Rowedder 1979; Hopkins and Torres 1989; Channichthyidae, ten species of Nototheniidae, and Lancraft et al. 1989, 1991; Duhamel 1998). Over the three species each of Rajidae and Zoarcidae. Common continental slope and shelf, notothenioids dominate the species were Trematomus eulepidotus (36.5%), T. scotti ichthyofauna (DeWitt 1970). Most members of this (32.0%), Prionodraco evansii (4.9%), T. -

Age and Growth of Brauer's Lanternfish Gymnoscopelus Braueri and Rhombic Lanternfish Krefftichthys Anderssoni (Family Myctophidae) in the Scotia Sea, Southern Ocean
Received: 22 July 2019 Accepted: 14 November 2019 DOI: 10.1111/jfb.14206 REGULAR PAPER FISH Age and growth of Brauer's lanternfish Gymnoscopelus braueri and rhombic lanternfish Krefftichthys anderssoni (Family Myctophidae) in the Scotia Sea, Southern Ocean Ryan A. Saunders1 | Silvia Lourenço2,3 | Rui P. Vieira4 | Martin A. Collins1 | Carlos A. Assis5,6 | Jose C. Xavier7,1 1British Antarctic Survey, Natural Environment Research Council, Cambridge, UK Abstract 2MARE - Marine and Environmental Sciences This study examines age and growth of Brauer's lanternfish Gymnoscopelus braueri and Centre, Polytechnic of Leiria, Peniche, Portugal rhombic lanternfish Krefftichthys anderssoni from the Scotia Sea in the Southern Ocean, 3CIIMAR/CIMAR, Interdisciplinary Centre of Marine and Environmental Research, through the analysis of annual growth increments deposited on sagittal otoliths. Oto- University of Porto, Matosinhos, Portugal lith pairs from 177 G. braueri and 118 K. anderssoni were collected in different seasons 4 Centre for Environment Fisheries and from the region between 2004 and 2009. Otolith-edge analysis suggested a seasonal Aquaculture Sciences, Lowestoft, UK change in opaque and hyaline depositions, indicative of an annual growth pattern, 5MARE-ULisboa – Marine and Environmental Sciences Centre, Faculty of Sciences, although variation within the populations of both species was apparent. Age estimates University of Lisbon, Lisbon, Portugal varied from 1 to 6 years for G. braueri (40 to 139 mm standard length; LS)andfrom 6Department of Animal Biology, Faculty of Sciences, University of Lisbon, Lisbon, Portugal 0 to 2 years for K. anderssoni (26 to 70 mm LS). Length-at-age data were broadly con- 7Department of Life Sciences, MARE-UC, sistent with population cohort parameters identified in concurrent length-frequency University of Coimbra, Coimbra, Portugal data from the region for both species. -

Diet and Trophic Niche of Antarctic Silverfish Pleuragramma
Journal of Fish Biology (2013) 82, 141–164 doi:10.1111/j.1095-8649.2012.03476.x, available online at wileyonlinelibrary.com Diet and trophic niche of Antarctic silverfish Pleuragramma antarcticum in the Ross Sea, Antarctica M. H. Pinkerton*, J. Forman, S. J. Bury, J. Brown, P. Horn and R. L. O’Driscoll National Institute of Water and Atmospheric Research Ltd, Private Bag 14901, Wellington 6241, New Zealand (Received 16 April 2012, Accepted 18 September 2012) The diet of Antarctic silverfish Pleuragramma antarcticum was evaluated by examining stomach ◦ ◦ ◦ ◦ contents of specimens collected in the Ross Sea (71 –77 S; 165 –180 E) in January to March 2008. Pleuragramma antarcticum (50–236 mm standard length, LS) and prey items were analysed for stable-isotopic composition of carbon and nitrogen. According to index of relative importance (IRI), which incorporates frequency of occurrence, mass and number of prey items, the most impor- tant prey items were copepods (81%IRI over all specimens), predominantly Metridia gerlachei and Paraeuchaeta sp., with krill and fishes having low IRI (2·2and5·6%IRI overall). According to mass of prey (M) in stomachs, however, fishes (P. antarcticum and myctophids) and krill domi- nated overall diet (48 and 22%M, respectively), with copepods being a relatively minor constituent of overall diet by mass (9·9%M). Piscivory by P. antarcticum occurred mainly in the extreme south- west of the region and near the continental slope. Krill identified to species level in P. antarcticum stomachs were predominantly Euphausia superba (14·1%M) with some Euphausia crystallopho- rias (4·8%M). -

Antartic Peninsula and Tierra Del Fuego: 100
ANTARCTIC PENINSULA & TIERRA DEL FUEGO BALKEMA – Proceedings and Monographs in Engineering, Water and Earth Sciences Antarctic Peninsula & Tierra del Fuego: 100 years of Swedish-Argentine scientific cooperation at the end of the world Edited by Jorge Rabassa & María Laura Borla Proceedings of “Otto Nordenskjöld’s Antarctic Expedition of 1901–1903 and Swedish Scientists in Patagonia: A Symposium”, held in Buenos Aires, La Plata and Ushuaia, Argentina, March 2–7, 2003. LONDON / LEIDEN / NEW YORK / PHILADELPHIA / SINGAPORE Cover photo information: “The Otto Nordenskjöld’s Expedition to Antarctic Peninsula, 1901–1903. The wintering party in front of the hut on Snow Hill, Antarctica, 30th September 1902. From left to right: Bodman, Jonassen, Nordenskjöld, Ekelöf, Åkerlund and Sobral. Photo: G. Bodman. From the book: Otto Nordenskjöld & John Gunnar Andersson, et al., “Antarctica: or, Two Years amongst the Ice of the South Pole” (London: Hurst & Blackett., 1905)”. Taylor & Francis is an imprint of the Taylor & Francis Group, an informa business This edition published in the Taylor & Francis e-Library, 2007. “To purchase your own copy of this or any of Taylor & Francis or Routledge’s collection of thousands of eBooks please go to www.eBookstore.tandf.co.uk.” © 2007 Taylor & Francis Group plc, London, UK All rights reserved. No part of this publication or the information contained herein may be reproduced, stored in a retrieval system, or transmitted in any form or by any means, electronic, mechanical, by photocopying, recording or otherwise, without written prior permission from the publishers. Although all care is taken to ensure the integrity and quality of this publication and the information herein, no responsibility is assumed by the publishers nor the author for any damage to property or persons as a result of operation or use of this publication and/or the information contained herein. -

Main Drivers of Mercury Levels in Southern Ocean Lantern Fish Myctophidae
Main drivers of mercury levels in Southern Ocean Lantern fish Myctophidae José Seco, José Xavier, Paco Bustamante, Joao Coelho, Ryan Saunders, Nicole Ferreira, Sophie Fielding, Miguel Pardal, Gabriele Stowasser, Thainara Viana, et al. To cite this version: José Seco, José Xavier, Paco Bustamante, Joao Coelho, Ryan Saunders, et al.. Main drivers of mercury levels in Southern Ocean Lantern fish Myctophidae. Environmental Pollution, Elsevier, 2020, 264C (114711), 10.1016/j.envpol.2020.114711. hal-02570100 HAL Id: hal-02570100 https://hal.archives-ouvertes.fr/hal-02570100 Submitted on 11 May 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Main drivers of mercury levels in Southern Ocean Lantern fish Myctophidae José Seco*1,2, José C. Xavier3,4, Paco Bustamante5, João P. Coelho6, Ryan A. Saunders3, Nicole Ferreira1, Sophie Fielding3, Miguel A. Pardal8, Gabriele Stowasser3, Thainara Viana1, Geraint A. Tarling3, Eduarda Pereira1, Andrew S. Brierley2 1 Department of Chemistry and CESAM/REQUIMTE, University of Aveiro, 3810-193 Aveiro, Portugal 2 Pelagic -
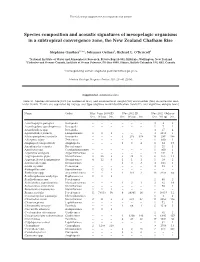
Marine Ecology Progress Series 503:23
The following supplement accompanies the article 1 Species composition and acoustic signatures of mesopelagic organisms in a subtropical convergence zone, the New Zealand Chatham Rise Stéphane Gauthier1, 2,*, Johannes Oeffner1, Richard L. O’Driscoll1 1National Institute of Water and Atmospheric Research, Private Bag 14-901, Kilbirnie, Wellington, New Zealand 2Fisheries and Oceans Canada, Institute of Ocean Sciences, PO Box 6000, Sidney, British Columbia V8L 4B2, Canada *Corresponding author: [email protected] Marine Ecology Progress Series: 503: 23–40 (2014) Supplement. Additional data Table S1. Species occurrence (Occ.) in number of tows, and abundance in weight (Wt) and number (No.) in successful mid - water trawls. Trawls are separated by voyage and type (daytime mark identification trawls ID, and nighttime oblique tows) Name Order May−June 2008 ID Nov 2011 ID Nov 2011 Oblique Occ. Wt (g) No. Occ. Wt (g) No. Occ. Wt (g) No. Acanthephyra pelagica Decapoda − − − − − − 3 9 3 Acanthephyra quadrispinosa Decapoda − − − − − − 1 7 1 Acanthephyra spp. Decapoda − − − − − − 3 17 4 Agrostichthys parkeri Lampriformes 1 8 1 − − − 3 1813 3 Alainopasiphaea australis Decapoda − − − 3 273 356 14 287 366 Allocyttus niger Zeiformes − − − − − − 1 600 1 Amphipod (unspecified) Amphipoda − − − 1 3 4 8 34 17 Anoplogaster cornuta Beryciformes − − − − − − 1 25 1 Apristurus spp. Carcharhiniformes − − − − − − 1 900 1 Argentina elongata Argentiniformes − − − − − − 4 541 6 Argyropelecus gigas Stomiiformes 2 20 2 2 38 5 8 165 12 Argyropelecus hemigymnus Stomiiformes 4 12 6 1 1 1 1 19 2 Astronesthes spp. Stomiiformes − − − 1 3 3 3 103 3 Atolla wyvillei Coronatae − − − − − − 4 83 7 Bathophilus ater Stomiiformes 1 12 1 − − − − − − Bathylagus spp. -

Marine Ecology Progress Series 370:239
Vol. 370: 239–247, 2008 MARINE ECOLOGY PROGRESS SERIES Published October 28 doi: 10.3354/meps07673 Mar Ecol Prog Ser Stable isotopes reveal the trophic position and mesopelagic fish diet of female southern elephant seals breeding on the Kerguelen Islands Y. Cherel1,*, S. Ducatez1, C. Fontaine1, P. Richard2, C. Guinet1 1Centre d’Etudes Biologiques de Chizé, UPR 1934 du CNRS, BP 14, 79360 Villiers-en-Bois, France 2Centre de Recherche sur les Ecosystèmes Littoraux Anthropisés, UMR 6217 du CNRS-IFREMER-ULR, Place du Séminaire, BP 5, 17137 L’Houmeau, France ABSTRACT: Trophic interactions between organisms are the main drivers of ecosystem dynamics, but scant dietary information is available for wide-ranging predators during migration. We investi- gated feeding habits of a key consumer of the Southern Ocean, the southern elephant seal Miroun- gia leonina, by comparing its blood δ13C and δ15N values with those of various marine organisms, including crustaceans, squid, fishes, seabirds and fur seals. At the end of winter, δ13C values (–23.1 to –20.1‰) indicate that female elephant seals forage mainly in the vicinity of the Polar Front and in the Polar Frontal Zone. Trophic levels derived from δ15N values (trophic level = 4.6) show that the southern elephant seal is a top consumer in the pelagic ecosystem that is dominated by colossal squid. The mean δ15N value of seals (10.1 ± 0.3‰) indicates that they are not crustacean eaters, but instead feed on crustacean-eating prey. Surprisingly, most of the previously identified prey species have isotope δ13C and δ15N values that do not fit with those of potential food items. -

Company Vendor ID (Decimal Format) (AVL) Ditest Fahrzeugdiagnose Gmbh 4621 @Pos.Com 3765 0XF8 Limited 10737 1MORE INC
Vendor ID Company (Decimal Format) (AVL) DiTEST Fahrzeugdiagnose GmbH 4621 @pos.com 3765 0XF8 Limited 10737 1MORE INC. 12048 360fly, Inc. 11161 3C TEK CORP. 9397 3D Imaging & Simulations Corp. (3DISC) 11190 3D Systems Corporation 10632 3DRUDDER 11770 3eYamaichi Electronics Co., Ltd. 8709 3M Cogent, Inc. 7717 3M Scott 8463 3T B.V. 11721 4iiii Innovations Inc. 10009 4Links Limited 10728 4MOD Technology 10244 64seconds, Inc. 12215 77 Elektronika Kft. 11175 89 North, Inc. 12070 Shenzhen 8Bitdo Tech Co., Ltd. 11720 90meter Solutions, Inc. 12086 A‐FOUR TECH CO., LTD. 2522 A‐One Co., Ltd. 10116 A‐Tec Subsystem, Inc. 2164 A‐VEKT K.K. 11459 A. Eberle GmbH & Co. KG 6910 a.tron3d GmbH 9965 A&T Corporation 11849 Aaronia AG 12146 abatec group AG 10371 ABB India Limited 11250 ABILITY ENTERPRISE CO., LTD. 5145 Abionic SA 12412 AbleNet Inc. 8262 Ableton AG 10626 ABOV Semiconductor Co., Ltd. 6697 Absolute USA 10972 AcBel Polytech Inc. 12335 Access Network Technology Limited 10568 ACCUCOMM, INC. 10219 Accumetrics Associates, Inc. 10392 Accusys, Inc. 5055 Ace Karaoke Corp. 8799 ACELLA 8758 Acer, Inc. 1282 Aces Electronics Co., Ltd. 7347 Aclima Inc. 10273 ACON, Advanced‐Connectek, Inc. 1314 Acoustic Arc Technology Holding Limited 12353 ACR Braendli & Voegeli AG 11152 Acromag Inc. 9855 Acroname Inc. 9471 Action Industries (M) SDN BHD 11715 Action Star Technology Co., Ltd. 2101 Actions Microelectronics Co., Ltd. 7649 Actions Semiconductor Co., Ltd. 4310 Active Mind Technology 10505 Qorvo, Inc 11744 Activision 5168 Acute Technology Inc. 10876 Adam Tech 5437 Adapt‐IP Company 10990 Adaptertek Technology Co., Ltd. 11329 ADATA Technology Co., Ltd. -

Sent to the Governor
HOUSE JOURNAL EIGHTY-THIRD LEGISLATURE, REGULAR SESSION PROCEEDINGS EIGHTY-THIRD DAY Ð MONDAY, MAY 27, 2013 The house met at 10 a.m. and was called to order by the speaker. The roll of the house was called and a quorum was announced present (Recordi1374). Present Ð Mr. Speaker; Allen; Alonzo; Alvarado; Anderson; Ashby; Aycock; Bell; Bohac; Bonnen, D.; Bonnen, G.; Branch; Burkett; Burnam; Button; Callegari; Canales; Capriglione; Carter; Clardy; Coleman; Collier; Cook; Cortez; Craddick; Creighton; Crownover; Dale; Darby; Davis, J.; Davis, S.; Davis, Y.; Deshotel; Dukes; Dutton; Eiland; Elkins; Fallon; Farias; Farney; Farrar; Fletcher; Flynn; Frank; Frullo; Geren; Giddings; Goldman; Gonzales; GonzaÂlez, M.; Gonzalez, N.; Gooden; Guerra; Guillen; Gutierrez; Harless; Harper-Brown; Hernandez Luna; Herrero; Hilderbran; Howard; Huberty; Hughes; Hunter; Isaac; Johnson; Kacal; Keffer; King, K.; King, P.; King, S.; King, T.; Kleinschmidt; Klick; Kolkhorst; Krause; Kuempel; Larson; Laubenberg; Lavender; Leach; Lewis; Longoria; Lozano; Lucio; MaÂrquez; Martinez; Martinez Fischer; McClendon; MeneÂndez; Miles; Miller, D.; Miller, R.; Moody; Morrison; MunÄoz; Murphy; Naishtat; NevaÂrez; Oliveira; Orr; Otto; Paddie; Parker; Patrick; Perez; Perry; Phillips; Pickett; Pitts; Price; Raney; Ratliff; Raymond; Reynolds; Riddle; Ritter; Rodriguez, E.; Rodriguez, J.; Rose; Sanford; Schaefer; Sheets; Sheffield, J.; Sheffield, R.; Simmons; Simpson; Smith; Smithee; Springer; Stephenson; Stickland; Strama; Taylor; Thompson, E.; Thompson, S.; Toth; Turner, C.; Turner, E.S.; Turner, S.; Villalba; Villarreal; Vo; Walle; White; Workman; Wu; Zedler; Zerwas. Absent Ð Anchia. The speaker recognized Representative Larson who introduced Reverend Trey H. Little, senior pastor, First Presbyterian Church, San Antonio, who offered the invocation as follows: Gracious and loving God, in this moment of quietness, we pause not to escape the realities of our day, but instead to acknowledge that being "one nation under God" is part of the fabric of our society. -

Atratus (Camarodonta: Echinometridae) After Eight Decades from the Rocky Shore of Kodiyaghat (Port Blair), South Andaman Islands, India
OPEN ACCESS All arfcles publfshed fn the Journal of Threatened Taxa are regfstered under Creafve Commons Atrfbufon 4.0 Interna - fonal Lfcense unless otherwfse menfoned. JoTT allows unrestrfcted use of arfcles fn any medfum, reproducfon and dfstrfbufon by provfdfng adequate credft to the authors and the source of publfcafon. Journal of Threatened Taxa The fnternafonal journal of conservafon and taxonomy www.threatenedtaxa.org ISSN 0974-7907 (Onlfne) | ISSN 0974-7893 (Prfnt) Note Reappearance of the rare Shfngle Urchfn Colobocentrotus (Podophora ) atratus (Camarodonta : Echfnometrfdae ) after efght decades from the rocky shore of Kodfyaghat (Port Blafr), South Andaman Islands, Indfa Vfkas Pandey & T. Ganesh 26 September 2016 | Vol. 8 | No. 11 | Pp. 9377–9380 10.11609/ jot. 2406 .8.1 1.9377-9380 For Focus, Scope, Afms, Polfcfes and Gufdelfnes vfsft htp://threatenedtaxa.org/About_JoTT.asp For Arfcle Submfssfon Gufdelfnes vfsft htp://threatenedtaxa.org/Submfssfon_Gufdelfnes.asp For Polfcfes agafnst Scfenffc Mfsconduct vfsft htp://threatenedtaxa.org/JoTT_Polfcy_agafnst_Scfenffc_Mfsconduct.asp For reprfnts contact <[email protected]> Publfsher/Host Partner Threatened Taxa Journal of Threatened Taxa | www.threatenedtaxa.org | 26 September 2016 | 8(11): 9377–9380 Note Reappearance of the rare Shingle Urchin a substantial group of marine Colobocentrotus (Podophora) atratus organisms including sea cucumbers, (Camarodonta: Echinometridae) after sea stars, brittle stars, sea urchins, eight decades from the rocky shore of and feather stars reach highest ISSN 0974-7907 (Online) diversity in reef environments, but ISSN 0974-7893 (Print) Kodiyaghat (Port Blair), South Andaman are also widespread on shallow Islands, India shores. Due to their wide trophic OPEN ACCESS preferences as, detritivores, filter- Vikas Pandey 1 & T. -

Size, Age, Growth and Mortality of the Green Sea Urchin Strongylocentrotus Droebachiensis in Vega and Hammerfest
Size, age, growth and mortality of the green sea urchin Strongylocentrotus droebachiensis in Vega and Hammerfest, Norway. Sandra Gran Stadniczeñko Master thesis (2011) Program for Marine Biology and Limnology Department of biology University of Oslo ACKNOWLEDGMENTS In order to make this thesis in the best possible way it was necessary to have the support of many people whom I want to thank. First of all, the best of thanks to my supervisors Stein Fredriksen1 and Kjell Magnus Norderhaug2 for their support and for giving me the opportunity to be a part of the team. I will not forget to also thank Hartvig Christie2 and Camilla With Fagerli2 for helping me with my doubts during this period. Immense thanks to Karl Inne Ugland1 for all the help when I got lost in the complex world of statistics. A more than special thanks to my boyfriend Johann. You have been without doubt the best support I've had in the difficult last months. A special thanks is not enough to thank you for all your patience and support, and for taking care of me during this time. Thanks a lot to all of you who have been next to me since childhood, and college period. To my brother Timur and friends Rocío, Diana, Isabel, Tania, Paloma and FranLu. Even though you have been so far from me those last two years, your support and friendship have helped me going on through the tough moments away from home. Thanks to you too, Elisabeth, Fidel and Javi. I have only known you for this last years but you have always been there when I needed your help and support. -
Echinodermata: Echinoidea)
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by PubMed Central Evolution of a Novel Muscle Design in Sea Urchins (Echinodermata: Echinoidea) Alexander Ziegler1*, Leif Schro¨ der2, Malte Ogurreck3, Cornelius Faber4, Thomas Stach5 1 Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts, United States of America, 2 Molecular Imaging Group, Leibniz-Institut fu¨r Molekulare Pharmakologie, Berlin, Germany, 3 Institut fu¨r Werkstoffforschung, Helmholtz-Zentrum Geesthacht, Geesthacht, Germany, 4 Institut fu¨r Klinische Radiologie, Universita¨tsklinikum Mu¨nster, Mu¨nster, Germany, 5 Institut fu¨r Biologie, Freie Universita¨t Berlin, Berlin, Germany Abstract The sea urchin (Echinodermata: Echinoidea) masticatory apparatus, or Aristotle’s lantern, is a complex structure composed of numerous hard and soft components. The lantern is powered by various paired and unpaired muscle groups. We describe how one set of these muscles, the lantern protractor muscles, has evolved a specialized morphology. This morphology is characterized by the formation of adaxially-facing lobes perpendicular to the main orientation of the muscle, giving the protractor a frilled aspect in horizontal section. Histological and ultrastructural analyses show that the microstructure of frilled muscles is largely identical to that of conventional, flat muscles. Measurements of muscle dimensions in equally-sized specimens demonstrate that the frilled muscle design, in comparison to that of the flat muscle type, considerably increases muscle volume as well as the muscle’s surface directed towards the interradial cavity, a compartment of the peripharyngeal coelom. Scanning electron microscopical observations reveal that the insertions of frilled and flat protractor muscles result in characteristic muscle scars on the stereom, reflecting the shapes of individual muscles.