Specific Richness, Size Classes and Growth Parameters of the Main Fish Species in the Upper Mouhoun River Basin in Burkina Faso
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

§4-71-6.5 LIST of CONDITIONALLY APPROVED ANIMALS November
§4-71-6.5 LIST OF CONDITIONALLY APPROVED ANIMALS November 28, 2006 SCIENTIFIC NAME COMMON NAME INVERTEBRATES PHYLUM Annelida CLASS Oligochaeta ORDER Plesiopora FAMILY Tubificidae Tubifex (all species in genus) worm, tubifex PHYLUM Arthropoda CLASS Crustacea ORDER Anostraca FAMILY Artemiidae Artemia (all species in genus) shrimp, brine ORDER Cladocera FAMILY Daphnidae Daphnia (all species in genus) flea, water ORDER Decapoda FAMILY Atelecyclidae Erimacrus isenbeckii crab, horsehair FAMILY Cancridae Cancer antennarius crab, California rock Cancer anthonyi crab, yellowstone Cancer borealis crab, Jonah Cancer magister crab, dungeness Cancer productus crab, rock (red) FAMILY Geryonidae Geryon affinis crab, golden FAMILY Lithodidae Paralithodes camtschatica crab, Alaskan king FAMILY Majidae Chionocetes bairdi crab, snow Chionocetes opilio crab, snow 1 CONDITIONAL ANIMAL LIST §4-71-6.5 SCIENTIFIC NAME COMMON NAME Chionocetes tanneri crab, snow FAMILY Nephropidae Homarus (all species in genus) lobster, true FAMILY Palaemonidae Macrobrachium lar shrimp, freshwater Macrobrachium rosenbergi prawn, giant long-legged FAMILY Palinuridae Jasus (all species in genus) crayfish, saltwater; lobster Panulirus argus lobster, Atlantic spiny Panulirus longipes femoristriga crayfish, saltwater Panulirus pencillatus lobster, spiny FAMILY Portunidae Callinectes sapidus crab, blue Scylla serrata crab, Samoan; serrate, swimming FAMILY Raninidae Ranina ranina crab, spanner; red frog, Hawaiian CLASS Insecta ORDER Coleoptera FAMILY Tenebrionidae Tenebrio molitor mealworm, -

Length-Weight Relationship and Condition Factor of Six Cichlid (Cichilidae: Perciformis) Species of Anambra River, Nigeria
Journal of Fisheries and Aquaculture ISSN: 0976-9927 & E-ISSN: 0976-9935, Volume 4, Issue 2, 2013, pp.-82-86. Available online at http://www.bioinfopublication.org/jouarchive.php?opt=&jouid=BPJ0000265 LENGTH-WEIGHT RELATIONSHIP AND CONDITION FACTOR OF SIX CICHLID (CICHILIDAE: PERCIFORMIS) SPECIES OF ANAMBRA RIVER, NIGERIA ATAMA C.I.1, OKEKE O.C.1, EKEH F.N.1, EZENWAJI N.E.1, ONAH I.E.1, IVOKE N.1, ONOJA U.S.2 AND EYO J.E.1* 1Department of Zoology and Environmental Biology, University of Nigeria, Nsukka, Enugu State, Nigeria. 2Department of Home Science, Nutrition and Dietetics, Faculty of Agriculture, University of Nigeria, Nsukka, Enugu State, Nigeria. *Corresponding Author: Email- [email protected] Received: June 10, 2013; Accepted: July 11, 2013 Abstract- The length-weight relationship and condition factor of six cichlid fish species inhabiting Anambra River were studied. The cichlid fish species were Chromidotilapia guntheri Sauvage 1882, Hemichromis bimaculatus Gill 1862, Tilapia zillii Gervais 1848, Hemichromis fasciatus Peters 1857, Tilapia mariae Boulenger 1899 and Oreochromis niloticus (Linnaeus 1758), obtained from fish landing sites of the river at Oguru- gu, Nsugbe and Otuocha. C. guntheri, H. bimaculatus and T. zillii exhibited positive allometric growth with b = 3.452, 3.828 and 3.210, respec- tively, while H. fasciatus, T. mariae and O. niloticus exhibited negative allometric growth with b = 2.667, 2.272 and 2.792, respectively. There was difference in the condition factors for the combined fish species and the monthly factor for each fish species studied: C. guntheri was 3.44 ± 0.39, H. -

Food and Feeding Habits of Tilapia Zilli (Pisces: Cichlidae) in River Otamiri South-Eastern Nigeria
Bioscience Discovery 3(2):146-148, June 2012 ISSN: 2229-3469 (Print) FOOD AND FEEDING HABITS OF TILAPIA ZILLI (PISCES: CICHLIDAE) IN RIVER OTAMIRI SOUTH-EASTERN NIGERIA Agbabiaka L. A. Department of Fisheries Technology, Federal Polytechnic Nekede, Owerri, Nigeria P.M.B. 1036, OWERRI, IMO STATE. NIGERIA [email protected] ABSTRACT Dietary habits of Tilapia zilli (Gervais, 1848) was studied in River Otamiri, Imo State, Nigeria. Fish specimens were procured from Artisanal fishermen every two weeks. Specimens were usually injected 4% formalin at the fishing station prior to laboratory analysis. A total of 97 specimens were analyzed for gut contents using Numerical and frequency of occurrence methods. Data collected showed that Tilapia zilli is an Omnivorous fish with dietary preference for Algae (71.05% and 59.52%), vegetative matter (10.52% and 50.00%), detritus (0% and 11.90%) and aquatic invertebrates larvae such as Chaoborus larvae (52.63% and 47.61%) and Chironomid larvae (31.58% and 21.43%) for juveniles and adult Tilapia respectively. Key words: Dietary habits, Tilapia zilli, River Otamiri, Omnivorous. INTRODUCTION MATERIALS AND METHODS Family Cichlidae comprising of Tilapia and River Otamiri lies between latitude 50 301 and 70 Hemichromis species are endemic to Nigeria, it is 301 North, and longitude 50 390 and 50 421 East. The widely distributed in Nigeria waters and second entire study area is about 20km representing the most abundant fish species at River Otamiri Southern part of the River along Obinze-Umuagwo (Agbabiaka, 2010). Various researchers have stretch in Imo State, Nigeria. Three sampling points investigated food and feeding habits of Cichlids and were located namely Obinze, Mgbirichi, and other commercially important fishes in Nigeria and Umuagwo which were about 7km intervals. -

DNA Barcoding Discriminates Freshwater Fishes from Southeastern Nigeria and Provides River System-Level Phylogeographic Resoluti
Mitochondrial DNA, 2011; Early Online: 1–9 DNA barcoding discriminates freshwater fishes from southeastern Nigeria and provides river system-level phylogeographic resolution within some species CHRISTOPHER D. NWANIa, SVEN BECKERb, HEATHER E. BRAIDb, EMMANUEL F. UDEc, OKECHUKWU I. OKOGWUa, & ROBERT HANNERb aDepartment of Applied Biology, Ebonyi State University, Abakaliki, Nigeria, bDepartment of Integrative Biology, Biodiversity Institute of Ontario, University of Guelph, Guelph, Ontario, Canada, and cFisheries and Aquaculture, Ebonyi State University, Abakaliki, Nigeria (Received 17 August 2010; revised 28 October 2010; accepted 28 October 2010) Abstract Background and aims: Fishes are the main animal protein source for human beings and play a vital role in aquatic ecosystems and food webs. Fish identification can be challenging, especially in the tropics (due to high diversity), and this is particularly true for larval forms or fragmentary remains. DNA barcoding, which uses the 50 region of the mitochondrial cytochrome c oxidase subunit I (cox1) as a target gene, is an efficient method for standardized species-level identification for biodiversity assessment and conservation, pending the establishment of reference sequence libraries. Materials and methods: In this study, fishes were collected from three rivers in southeastern Nigeria, identified morphologically, and imaged digitally. DNA was extracted, PCR-amplified, and the standard barcode region was bidirectionally sequenced for 363 individuals belonging to 70 species in 38 genera. All specimen provenance data and associated sequence information were For personal use only. recorded in the barcode of life data systems (BOLD; www.barcodinglife.org). Analytical tools on BOLD were used to assess the performance of barcoding to identify species. Results: Using neighbor-joining distance comparison, the average genetic distance was 60-fold higher between species than within species, as pairwise genetic distance estimates averaged 10.29% among congeners and only 0.17% among conspecifics. -

Yaoure ESIA Appendix 18 Critical Habitat Assessment
Appendix 18 Critical Habitat Assessment Yaoure Gold Project, Côte d’Ivoire Submitted to Amara Mining Submitted By Amec Foster Wheeler Earth & Environmental (UK) Ltd. ESIA REPORT, YAOURE GOLD PROJECT APPENDIX 18 CRITICAL HABITAT ASSESSMENT JUNE 2015 R E P O R T I S S U E F ORM Client Name Amara Mining plc Project Name Yaoure Gold Project Report Title Critical Habitat Assessment Document Status Final Issue No. 1 Issue Date 12 November 2015 Report Number Document Reference 7879140169 A169-15-R2325 Author Dr. Genevieve Campbell Signature & 24 July 2015 Reviewer Dr. Christian Kunze Signature & 24 July 2015 Project Manager Approval Dr. Christian Kunze 12 November 2015 D ISCLAIMER This report was prepared exclusively for the client above by Amec Foster Wheeler Earth & Environmental (UK) Ltd. (Amec Foster Wheeler). The quality of information, conclusions and estimates contained herein are consistent with the level of effort involved in Amec Foster Wheeler’s services and based on: i) information available at the time of preparation, ii) data supplied by outside sources and iii) the assumptions, conditions and qualifications set forth in this report. This report is intended for use by the above client subject to the terms and conditions of its contract with Amec Foster Wheeler. Any other use of, or reliance on, this report by any third party is at that party’s sole risk. Project No.: 7879140169 Report Issue Form ESIA REPORT, YAOURE GOLD PROJECT APPENDIX 18 CRITICAL HABITAT ASSESSMENT JUNE 2015 E X E C U T I V E S UMMARY This Critical Habitat Assessment aimed at evaluating Critical Habitat qualifying features present in the Project area in order to determine if they trigger Critical Habitat according to IFC PS6. -

Summary of Temperature Metrics for Aquatic Invasive Fish Species in the Prairie Region
Summary of Temperature Metrics for Aquatic Invasive Fish Species in the Prairie Region Theresa E. Mackey, Caleb T. Hasler, and Eva C. Enders Fisheries and Oceans Canada Ecosystems and Oceans Science Central and Arctic Region Freshwater Institute Winnipeg, MB R3T 2N6 2019 Canadian Technical Report of Fisheries and Aquatic Sciences 3308 1 Canadian Technical Report of Fisheries and Aquatic Sciences Technical reports contain scientific and technical information that contributes to existing knowledge but which is not normally appropriate for primary literature. Technical reports are directed primarily toward a worldwide audience and have an international distribution. No restriction is placed on subject matter and the series reflects the broad interests and policies of Fisheries and Oceans Canada, namely, fisheries and aquatic sciences. Technical reports may be cited as full publications. The correct citation appears above the abstract of each report. Each report is abstracted in the data base Aquatic Sciences and Fisheries Abstracts. Technical reports are produced regionally but are numbered nationally. Requests for individual reports will be filled by the issuing establishment listed on the front cover and title page. Numbers 1-456 in this series were issued as Technical Reports of the Fisheries Research Board of Canada. Numbers 457-714 were issued as Department of the Environment, Fisheries and Marine Service, Research and Development Directorate Technical Reports. Numbers 715-924 were issued as Department of Fisheries and Environment, Fisheries and Marine Service Technical Reports. The current series name was changed with report number 925. Rapport technique canadien des sciences halieutiques et aquatiques Les rapports techniques contiennent des renseignements scientifiques et techniques qui constituent une contribution aux connaissances actuelles, mais qui ne sont pas normalement appropriés pour la publication dans un journal scientifique. -

La Biodiversité En Côte D'ivoire : Etat Des Lieux Et Facteurs De Menace
La biodiversité en Côte d’Ivoire : Etat des lieux et facteurs de menace 5 Biodiversity of Côte d’Ivoire: Current state and threats L’une des caractéristiques les plus fascinantes de la nature est la diversité des êtres vivants dont elle regorge. La biodiversité est définit simplement comme la diversité de toutes les formes du vivant, incluant la diversité génétique, spécifique et écosystémique. Elle constitue la toile de la vie terrestre et aquatique dont nous sommes partie intégrante et dont nous dépendons totalement pour notre bien être et notre survie. Elle atteint son maximum dans les régions tropicales dont les forêts abritent plus de 50 % de la diversité biologique mondiale. En Côte d’Ivoire, la conservation de la biodiversité est une question majeure au regard de son importance et des menaces fréquentes qui pèsent sur elle. En effet, de par la diversité et la particularité de sa flore et de sa faune, la Côte d’Ivoire fait partie des zones prioritaires de conservationhot ( spot de biodiversité) en Afrique de l’Ouest. Cependant cette diversité biologique est fortement menacée par de nombreux facteurs dont les plus importants sont la déforestation, l’agriculture non durable, le braconnage, la pollution et les changements climatiques. Ces menaces qui affectent considérablement aussi bien les écosystè- mes que les organismes vivants qu’ils abritent, ont un effet déterminant sur l’économie et la qualité de la vie humaine. Ce chapitre a pour objectif de faire un état des lieux de la biodiversité en Côte d’Ivoire et d’identifier les facteurs pouvant menacer sa conservation. Il comprend trois parties qui s’articulent autour des composantes essentielles de la biodiversité. -

View/Download
CICHLIFORMES: Cichlidae (part 2) · 1 The ETYFish Project © Christopher Scharpf and Kenneth J. Lazara COMMENTS: v. 4.0 - 30 April 2021 Order CICHLIFORMES (part 2 of 8) Family CICHLIDAE Cichlids (part 2 of 7) Subfamily Pseudocrenilabrinae African Cichlids (Abactochromis through Greenwoodochromis) Abactochromis Oliver & Arnegard 2010 abactus, driven away, banished or expelled, referring to both the solitary, wandering and apparently non-territorial habits of living individuals, and to the authors’ removal of its one species from Melanochromis, the genus in which it was originally described, where it mistakenly remained for 75 years; chromis, a name dating to Aristotle, possibly derived from chroemo (to neigh), referring to a drum (Sciaenidae) and its ability to make noise, later expanded to embrace cichlids, damselfishes, dottybacks and wrasses (all perch-like fishes once thought to be related), often used in the names of African cichlid genera following Chromis (now Oreochromis) mossambicus Peters 1852 Abactochromis labrosus (Trewavas 1935) thick-lipped, referring to lips produced into pointed lobes Allochromis Greenwood 1980 allos, different or strange, referring to unusual tooth shape and dental pattern, and to its lepidophagous habits; chromis, a name dating to Aristotle, possibly derived from chroemo (to neigh), referring to a drum (Sciaenidae) and its ability to make noise, later expanded to embrace cichlids, damselfishes, dottybacks and wrasses (all perch-like fishes once thought to be related), often used in the names of African cichlid genera following Chromis (now Oreochromis) mossambicus Peters 1852 Allochromis welcommei (Greenwood 1966) in honor of Robin Welcomme, fisheries biologist, East African Freshwater Fisheries Research Organization (Jinja, Uganda), who collected type and supplied ecological and other data Alticorpus Stauffer & McKaye 1988 altus, deep; corpus, body, referring to relatively deep body of all species Alticorpus geoffreyi Snoeks & Walapa 2004 in honor of British carcinologist, ecologist and ichthyologist Geoffrey Fryer (b. -

The Reproductive Biology of the Fishes of Owa Stream, South - West Nigeria
Research Journal of Fisheries and Hydrobiology, 5(2): 81-84, 2010 © 2010, INSInet Publication The Reproductive Biology of the Fishes of Owa Stream, South - West Nigeria. K.B. Olurin and O.I. Odeyemi Department of Plant Science & Applied Zoology, Olabisi Onabanjo University, P.M.B. 2002, Ago-Iwoye, Nigeria. Abstract: Sex ratio, size at maturity, maturity stages, egg diameter and fecundity of some of the fishes of Owa stream, south – west Nigeria were investigated. Only Hemichromis fasciatus differed significantly from the 1:1 sex ratio. Size at maturity for females was 9.05cm for Chromidotilapia guntheri, 10.45cm for Tilapia mariae, and 8.3cm for Hemichromis fasciatus.Mature males were observed for specimens of C.guntheri, while the others had no mature males. The gonado-somatic indices increased with gonadal stage of development, and the values for males were lower than those of the females of corresponding stage of development. Egg diameter was highest in Xenomystus nigri, and least in Ctenoporna kingsleyae. There was a low correlation between fecundity and body weight in C. guntheri. Keywords: Owa stream, gonado – somatic index, size at maturity, sex ratio INTRODUCTION The specimens were sorted out in the laboratory and identified to species level using information Literature is replete with information on the provided by[8]. reproductive biology of the fishes of temperate streams The fishes were mopped on filter paper and their [e.g.,19,1,9,12], with a dearth of information on their total lengths and standard lengths were measured using tropical counterparts. a measuring board to the tenth of a centimeter. -

Ichthyological Exploration of Freshwaters an International Journal for Field-Orientated Ichthyology
Verlag Dr. Friedrich Pfeil ISSN 0936-9902 Excerpt from Ichthyological Exploration of Freshwaters An international journal for field-orientated ichthyology Volume 28 Number 2 This article may be used for research, teaching and private purposes. Exchange with other researchers is allowed on request only. Any substantial or systematic reproduction, re-distribution, re-selling in any form to anyone, in particular deposition in a library, institutional or private website, or ftp-site for public access, is expressly forbidden. Ichthyological Exploration of Freshwaters An international journal for field-orientated ichthyology Volume 28 • Number 2 • January 2018 pages 97-192, 40 figs., 14 tabs. Managing Editor Paulo H. F. Lucinda, Laboratório de Ictiologia Sistemática, Universidade Federal do Tocantins, P.O. Box 136, 77500-000 Porto Nacional, TO, Brazil E-mail [email protected] Honorary Editor Maurice Kottelat, Rue des Rauraques 6, CH-2800 Delémont, Switzerland Associate Editors Ralf Britz, Department of Zoology, The Natural History Museum, London, United Kingdom Kevin W. Conway, Department of Wildlife and Fisheries Sciences, Texas A&M University, College Station, USA Jörg Freyhof, Leibniz-Institute of Freshwater Ecology and Inland Fisheries, Berlin, Germany Sven O. Kullander, Naturhistoriska Riksmuseet, Stockholm, Sweden Heok Hee Ng, Lee Kong Chian Natural History Museum, National University of Singapore, Singapore Editorial Advisory Board Helen K. Larson, Museum and Art Gallery of the Northern Territory, Darwin, Australia Lukas Rüber, Naturhistorisches Museum, Bern, Switzerland Ivan Sazima, Museu de Zoologia, Unicamp, Campinas, Brazil Paul H. Skelton, South African Institute for Aquatic Biodiversity, Grahamstown, South Africa Tan Heok Hui, Lee Kong Chian Natural History Museum, National University of Singapore, Singapore Ichthyological Exploration of Freshwaters is published quarterly Subscriptions should be addressed to the Publisher: Verlag Dr. -
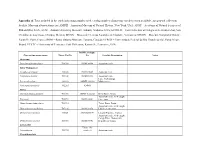
Appendix A. Taxa Included in the Study Indicating Samples Used, Catalog Number of Museum Vouchers When Available, and General Collection Locality
Appendix A. Taxa included in the study indicating samples used, catalog number of museum vouchers when available, and general collection locality. Museum abbreviations are: AMNH – American Museum of Natural History, New York, USA; ANSP – Academy of Natural Sciences of Philadelphia, USA; AUM – Auburn University Museum, Auburn, Alabama, USA; ECOSUR – Fish Collection at Colegio de la Frontera Sur, San Cristóbal de Las Casas, Chiapas, Mexico; MCNG – Museo de Ciencias Naturales de Guanare, Venezuela; MNHN – Muséum National d’Histoire Naturelle, Paris, France; ROM – Royal Ontario Museum, Toronto, Canada; UFRGS – Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil; UTFTC – University of Tennessee Fish Collection, Knoxville, Tennessee, USA. ROM Catalogue Current taxonomy name Tissue Cat No No Locality Description Notes Outgroup Pseudetroplus maculatus T14743 ROM 98998 Aquarium trade India-Madagascar Etroplus suratensis T13505 ROM 93809 Aquarium trade Paratilapia polleni T13100 ROM 88333 Aquarium trade Lake Andrapongy, Paretroplus damii 201936 AMNH 201936 Madagascar Paretroplus polyactis T12265 AMNH Africa Chromidotilapia guntheri T11700 AMNH I-226361 Beffa River, Benin Aquarium trade, wild caught, Etia nguti T10792 ROM 88042 Cameroon Hemichromis bimaculatus T11719 Tchan Duga, Benin Aquarium trade, wild caught, Heterochromis multidens T07136 ROM 88350 Lobeke, Cameroon Oreochromis niloticus 9092S AMNH254194 Littoral Province, Guinea Aquarium trade, wild caught, Congo River, Democratic Orthochromis stormsi T10766 ROM 88041 Republic of -

Mitochondrial Phylogeny and Phylogeography of East African
BMC Evolutionary Biology BioMed Central Research article Open Access Mitochondrial phylogeny and phylogeography of East African squeaker catfishes (Siluriformes: Synodontis) Stephan Koblmüller1, Christian Sturmbauer1, Erik Verheyen2, Axel Meyer3 and Walter Salzburger*3 Address: 1Department of Zoology, Karl-Franzens-University Graz, Universitätsplatz 2, 8010 Graz, Austria, 2Vertebrate Department, Royal Belgian Institute of Natural Sciences, 1000 Brussels, Belgium and 3Lehrstuhl für Zoologie und Evolutionsbiologie, Department of Biology, University of Konstanz, 78467 Konstanz, Germany Email: Stephan Koblmüller - [email protected]; Christian Sturmbauer - [email protected]; Erik Verheyen - [email protected]; Axel Meyer - [email protected]; Walter Salzburger* - walter.salzburger@uni- konstanz.de * Corresponding author Published: 19 June 2006 Received: 10 April 2006 Accepted: 19 June 2006 BMC Evolutionary Biology 2006, 6:49 doi:10.1186/1471-2148-6-49 This article is available from: http://www.biomedcentral.com/1471-2148/6/49 © 2006 Koblmüller et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Abstract Background: Squeaker catfishes (Pisces, Mochokidae, Synodontis) are widely distributed throughout Africa and inhabit a biogeographic range similar to that of the exceptionally diverse cichlid fishes, including the three East African Great Lakes and their surrounding rivers. Since squeaker catfishes also prefer the same types of habitats as many of the cichlid species, we hypothesized that the East African Synodontis species provide an excellent model group for comparative evolutionary and phylogeographic analyses.